Die Thermoplasmata sind fakultativ anaerob, d. h. sie können sowohl in Gegenwart als auch in Abwesenheit von freiem Sauerstoff leben.
Sie typischerweise thermoacidophil, d. h. sie benötigen meist hohe Temperaturen und einen niedrigen pH-Wert (acidophil = säureliebend).
Die meisten Mitglieder der Thermoplasmata sind thermophil.
Als acidophile Organismen wachsen die Thermoplasmata optimal bei einem pH-Wert unter 2.[4]
Es gibt jedoch auch einige nicht kultivierte Thermoplasmata-Mitglieder, die unter gemäßigten Bedingungen an der Meeresoberfläche und am Meeresboden leben.[5][6][7]
Zusammen mit einigen nahe verwandten Gruppen, insbesondere der Klasse Poseidoniia, bilden die Thermoplasmata eine Klade, die teilweise als Phylum Thermoplasmatota[8][9][10] innerhalb des ReichsMethanobacteriati (früher Superphylum Euryarchaeida oder Euryarchaeota sensu lato) angesehen wird.
Andere Taxonomien sehen die Thermoplasmata als Mitglied der Methanobacteriota (früher Phylum Euryarchaeota sensu stricto). Die Klasse Poseidoniia bildet mit einigen verwandten Gruppen dort das Phylum Poseidoniota.[2]
Die Archaeen der Gattung Thermoplasma leben beispielsweise in Halden von Kohleschlacken, wo chemoautotropheBakterienEisenpyrit zu Schwefliger Säure und Schwefelsäure oxidieren. Dies führt zu hohen Temperaturen und einem niedrigen pH-Wert, was die Bildung kleiner Kohlenstoffmoleküle begünstigt, die von diesen Organismen verstoffwechselt werden können.
Die dieser Ordnung zugehörige Gattung Picrophilus war zur Zeit ihrer Erstbeschreibung 1996 der acidophilste aller bekannten Organismen, sie wächst bei einem pH-Wert von 0,06 oder weniger.[4]
Die Thermoplasmata haben meist keine Zellwand. Picrophilus ist hierbei eine Ausnahme, ihre Zellmembran besteht aus großen Mengen von Lipopolysacchariden und Glykoproteinen, die sie vor Hitze und Säuren schützen.[4]
„Ca. Lutacidiplasmatales“
Die ersten Hinweise auf die ursprünglich so genannte Terrestrial Miscellaneous Euryarchaeota Group (TMEG), heute per Vorschlag „Ca. Lutacidiplasmatales“, fanden sich durch Metagenomik-Analysen von Proben aus südafrikanischen Goldminen. Weitere Analysen lieferten den Nachweis, dass sie eine eigenständige taxonomische Klade bilden.[11]
Auf der Grundlage des Genoms ihres Vertreters TMEG-bg1 aus tiefen anaeroben Torfschichten ließen sich Vorhersagen über ihren Stoffwechsel treffen.[11]
Demnach kann TMEG-bg1 Fettsäuren durch anaerobe Atmung oder durch syntrophische Wechselwirkungen mit Methanogenen oxidieren.[12]
Im Jahr 2022 beschrieben Sheridan et al. die Gruppe als Kandidaten-Ordnung „Ca. Lutacidiplasmatales“, wobei die Teile „Luti“ und „acidi“ auf die Häufigkeit in saurem Boden und „plasma“ auf die Verwandtschaft mit den Thermoplasmata (bzw. Thermoplasmatota) hinweisen.[11]
„Ca. Yaplasmales“
Ebenfalls im Jahr 2022 identifizierten Zhang et al. die Gruppe „Ca. Yaplasmales“ als weitere Kandidaten-Ordnung in dieser Verwandtschaft.[13]
Methanomassiliicoccales
Eine weitere Ordnung der Thermoplasmata ist Methanomassiliicoccales, deren Mitglieder an der Methanogenese beteiligt sind.
Habitat
Die Thermoplasmata kommen in zahlreichen Nischen und Umgebungen vor, die ersten identifizierten Vertreter fanden sich in südafrikanischen Goldminen, die „Ca. Lutacidiplasmatales“ umfassen häufig vorkommende Arten auf dem Festland.[11]
Es wird angenommen, dass ihr (letzter) gemeinsamer Vorfahr auch schon thermophil war, und dass mehrere Arten erst später und unabhängig voneinander, d. h. im Zug zunehmender Diversität mesophil wurden.
Aufgrund der Genome von „Ca. Lutacidiplasmatales“ sind aerobe Atmung, Glykolyse und Säuretoleranz in dieser Ordnung vermutlich wiederholt durch konvergente Evolution und horizontalen Gentransfer (HGT) entstanden.[11]
Andererseits entstanden auch polyextremophile Arten, mit Habitaten wie beispielsweise dem Großen Salzsee (Great Salt Lake, GSL) in den USA[14] oder den Natriumchloridseen der Kulundasteppe – wie z. B. T1Sed10_119m (Thermoplasmata PWHR01)[15] bei Мали́новое озероMalinowoje Osero, deutsch ‚Himbeersee‘[16] und CSSed10_214 (Thermoplasmata PWKY01)[17] im Kleinen Озеро ПетуховоOzero Petukhovo südöstlich von (СеверкаSeverka (Russland), an der Grenze zu Kasachstan.[18]
Die Thermoplasmata sind zur aerobenZellatmung fähig, es gibt strikt aerobe und fakultativ anaerobe Arten. Auf Basis der Sequenz von TMEG-bg1 (2022) wird angenommen, dass es mehrere anaeroben Arten gibt, die langkettige Fettsäuren und Sulfite abbauen und dabei Schwefelwasserstoff (H2S) produzieren.[11]
Vertreter der Ordnung „Ca. Yaplasmales“ sind am Kreislauf organischer Verbindungen in Tiefseesedimenten beteiligt und bauen Alkane mit einer Alkylsuccinat-Synthase ab. Von den drei Kladen A, B und C (den GTDB-Familien f__JAGMCJ01, f__JAJISG01 respektive f__RBG-16-68-12, s. u.) ist die Klade A basierend auf ihrem vollständigen Wood-Ljungdahl-Stoffwechselweg und ihren vermutlichen Genen für den Abbau von aromatischen und organischen Halogenverbindungen vermutlich mixotroph: Die Kladen B und C sind dagegen wahrscheinlich heterotroph, wobei sie Spermidin, Putrescin und aromatische Verbindungen abbauen können.[13]
Bedeutung für den Menschen
Arten der Methanomassiliicoccaceae zählen zu den häufigsten Vertretern im Verdauungstrakt, daneben kommen dort auch Arten der Methanomethylophilaceae vor. Diese beiden Gruppen treten dabei selten gemeinsam auf.[20]
Die genaue Taxonomie dieser Klasse ist noch in der Diskussion (Stand September 2025).
Die Ordnung „Ca. Aciduliprofundales“[22] [DVHE2 group][23] ist nach der NCBI-Taxonomie Mitglied des dortigen Phylums Thermoplasmatota, gehört aber nach der LPSN und der GTDB direkt zur Klasse Thermoplasmata.[3][22][23]
Die (in der LPSN mit Stand 24. August 2025 nicht gelistete) Ordnung „Ca. Lutacidiplasmatales“ gilt in diesen Taxonomien, darunter NCBI und Sheridan et al. (2022), als Mitglied des dortigen Phylums Thermoplasmatota.[9][11] Ihr Mitglied UBA184[11] residiert in der GTDB ebenfalls dort, und zwar in der GTDB-Ordnung o__UBA184 der Klasse Thermoplasmata. Dies impliziert, dass die GTDB-Ordnung o__UBA184 ein Synonym für „Ca. Lutacidiplasmatales“ ist.[24] UBA184 residiert auch in der NCBI-Taxonomie in der Klasse Thermoplasmata (allerdings dort ohne Ordnungszuweisung).[25] Der nahe verwandte Stamm AcS1-36[11] findet sich in derselben GTDB-Gattung. „Ca. Lutacidiplasmatales“ [o__UBA184] ist somit ein Mitglied der Klasse Thermoplasmata. Eine verwandte Klade (Ordnung) ist:[11]
Stamm SubAcS15-131, Ordnung Lunaplasmatales; gemäß NCBI ist der Stamm Mitglied der Klasse Thermoplasmata.[26][27][28] Diese Klade aus nur aus einer einzigen Spezies „Ca. Lunaplasma lacustris“ sollte nach Diamond et al. (2022) lediglich eine Familie innerhalb der von hnen vorgeschlagenen Ordnung „Ca. Angelarchaeales“ darstellen (s. u.).[19]
Die (in der LPSN mit Stand 24. August 2025 verwaiste[29]) Ordnung „Ca. Yaplasmales“ (früher als Klade RBG-16-68-12 bezeichnet) gilt nach Zheng et al. (2022) ebenfalls als Mitglied des Phylums Thermoplasmatota.[13] Ihr Mitglied, Archaeon bin162[13] residiert in der GTDB ebenfalls dort, und zwar in der GTDB-Ordnung o__RBG-16-68-12 der Klasse Thermoplasmata.[30] Verwandte Kladen (Ordnungen) sind nach Zheng et al. (2022):[13]
Stamm UBA10834, residiert in der GTDB in der GTDB-Ordnung o__UBA10834 [„Ca. Gimiplasmatales“, Diamond et al. (2022)[19]] innerhalb der Thermoplasmata.[31] Dieser und der Stamm RBG_19FT_COMBO_56_21 (GTDB-Spezies COMBO-56-21 sp001800815, GCA_001800815.1) residieren in der GTDB-Familie f__UBA10834. Zu dieser Ordnung gehört nach Hu et al. auch die Spezies „Ca. Gimiplasma haoranii“. Wegen vorausgesetzter Monophylie der vorgeschlagenen Taxa müssen daher „Ca. Gimiplasmataceae“ und f__UBA10834 synonym sein.[32]
Stamm SG8-5, residiert in der GTDB in der GTDB-Ordnung o__SG8-5 [„Ca. Proteinoplasmatales“, NCBI] innerhalb der Thermoplasmata[33][34]
„Ca. Angelarchaeales“, nahe verwandt mit Klade SG8-5, etwas weiter mit Klade UBA10834 und Methanomassiliicoccales [Methanomassillicoccales], enthält die Familie um „Ca. Lunaplasma lacustris“ SubAcS15-131.[19][35][36]
Als Mitglieder der Klasse Thermoplasmata gehören diese Beispiele in einer Taxonomie mit Phylum „Ca. Poseidoniota“ mit der Klasse dem Phylum Methanobacteriota an.
Liste
Die folgende Systematik hat den Stand 8. September 2025:
Stamm TMEG-bg1 alias bg1(S) oder Bg1 (Bg1 EMIRGE, T3M-75cm Bg1)[12] – Fundort: Metagenom von tiefen anoxischen Torfschichten(S) (S1 bog T3M), Marcell Experimental Forest im Chippewa National Forest, Minnesota, USA[12] Der Stamm ist offenbar identisch mit Stamm UBA61(N,G)[21] – Fundort: Metagenom einer Bodenprobe, Marcell Experimental Forest im Chippewa National Forest, Minnesota, USA(N)
Die folgenden Herleitungen stammen, wo nicht anders angegeben, aus der LPSN zum jeweiligen Taxon (ausgehend von Thermoplasmata):[2]
Die Bezeichnung der Typusgattung Thermoplasma leitet sich ab von altgriechischθέρμηthermē, deutsch ‚Hitze‘ und πλάσμαplásma, deutsch ‚formbare Masse‘, ‚etwas Geformtes‘, es wird damit also eine heiße oder Hitze liebende formbare Masse bezeichnet, was – wie bei den meisten anderen Vertretern der Thermoplasmatota – auf das Fehlen einer Zellwand hinweist.
Die Bezeichnung der Klasse Thermoplasmata leitet sich ab von der Typusgattung Thermoplasma, zusammen mit der Klassen kennzeichnenden Endung ‚-ata‘ (NominativPlural).
Die Bezeichnung des Phylums Thermoplasmatota leitet sich ab von der Typusklasse Thermoplasmata, zusammen mit der Phyla kennzeichnenden Endung ‚-ota‘.
Analog leitet sich die Bezeichnung der Ordnung Thermoplasmatales ebenfalls ab von der Typusklasse, aber mit der Ordnungen kennzeichnenden Endung ‚-ales‘.
Die Bezeichnung der Klasse Poseidoniia leitet sich ab von der Typusgattung Poseidonia, diese ist benannt nach Poseidon, dem griechischen Gott des Meeres; Poseidoniia („die Poseidonier“) bezeichnet also die Poseidonia-Klasse.
Die Bezeichnung der Ordnung Ca. Aciduliprofundales leitet sich her von ihrer Typusgattung Aciduliprofundum, diese setzt sich zusammen aus lateinischacidulus‚säuerlich‘ und profundus‚tief‘, angehängt ist der Suffix ‚-ales‘ für Ordnungen. Dies bezeichnet daher einen säureliebenden Organismus aus der Tiefe.
Die Bezeichnung der Ordnung Ca. Gimiplasmatales leitet sich ab von ihrer Typusgattung Gimiplasma, was sich zusammensetzt aus der Abkürzung GIM für das GuangdongInstitute of Microbiology und der Endung ‚-plasma‘ für eine formbare Masse; angehängt ist der Suffix ‚-ales‘ für Ordnungen.
Die Bezeichnung der Ordnung Ca. Halarchaeoplasmatales leitet sich ab von der Typusgattung Halarchaeoplasma, zusammengesetzt aus altgriechisch ᾰ̔́λςháls, deutsch ‚Salz‘, Genitiv ᾰ̔λόςhalos, dem Mittelteil ‚-archaeo-‘ zur Unterscheidung von Bakterien in ähnlichen Umgebungen, und der Endung ‚-plasma‘ für eine formbare Masse; angehängt ist der Suffix ‚-ales‘ für Ordnungen.
Die Bezeichnung der Ordnung (bzw. herabgestuft Familie) Ca. Lunaplasmatales leitet sich ab von ihrer Typusgattung Lunaplasma, diese ist zusammengesetzt aus lateinisch luna‚Mond‘ und der Endung ‚-plasma‘ für eine formbare Masse; angehängt ist der Suffix ‚-ales‘ für Ordnungen.
Die Bezeichnung der Ordnung Methanomassiliicoccales leitet sich ab von der Typusgattung Methanomassiliicoccus, zusammengesetzt aus den Namensteilen lateinisch ‚Methano-‘, Methan betreffend, ebenfalls lateinisch Massilia‚Marseille‘ und neulateinisch coccus von altgriechisch κόκκοςkókkos, deutsch ‚Kern, Korn‘; die Ordnung ist noch mit dem Ordnungs-Suffix ‚-ales‘ versehen. Die Bezeichnung steht also für methanbildende Kügelchen aus (gefunden oder erforscht in) Marseille.
Die Bezeichnung der Ordnung Ca. Sysuiplasmatales leitet sich ab von ihrer Typusgattung Sysuiplasma, deren Name kommt vom Akronym SYSU der Sun-Yat-sen-Universität (Guangzhou), wo diese Organismen erforscht wurden, und der Endung ‚-plasma‘ für eine formbare Masse; die Ordnung ist noch versehen mit dem entsprechenden Suffix ‚-ales‘.[50]
Die Bezeichnung der vorgeschlagenen Ordnung Ca. Yaplasmales leitet sich ab von chinesisch涯Ya, deutsch ‚Grenze‘, ‚-plasma-‘ für eine formbare Masse und dem Ordnungs-Suffix ‚-ales‘. Dies weist darauf hin, dass sie (unter anderem) in den nährstoffarmen Sedimenten der Tiefsee leben.[13]
Eine provisorische Bezeichnung für eine Reihe von Euryarchaeota-Kladen aus Tiefsee-Hydrothermalquell ist DHVE oder DHVEG, ausgeschrieben Deep-sea Hydrothermal Vent Euryarchaeota (group), gefolgt von einer Nummer.
Weiterführende Literatur
Wissenschaftliche Journale
Chao Lun Li, Yun Tao Jiang, Da Li Liu, JieLei Qian, Jing Ping Liang, Rong Shu: Prevalence and quantification of the uncommon Archaea phylotype Thermoplasmata in chronic periodontitis. In: Archives of Oral Biology. 59. Jahrgang, Nr.8, August 2014, S.822–828, doi:10.1016/j.archoralbio.2014.05.011, PMID 24859768 (englisch).
Morten Poulsen, Clarissa Schwab, Bent Borg Jensen, Ricarda M. Engberg, Anja Spang, Nuria Canibe, Ole Hojberg: Methylotrophic methanogenic Thermoplasmata implicated in reduced methane emissions from bovine rumen. In: Nature Communications. 4. Jahrgang, Nr.1428, Juni 2013, S.1947, doi:10.1038/ncomms2847, bibcode:2013NatCo...4.1947P (englisch).
Thomas Cavalier-Smith: The neomuran origin of archaebacteria, the negibacterial root of the universal tree and bacterial megaclassification. In: Int. J. Syst. Evol. Microbiol. 52. Jahrgang, Nr.1, 2002, S.7–76, doi:10.1099/00207713-52-1-7, PMID 11837318 (englisch).
Anna N. Rasmussen, Bradley B. Tolar, John R. Bargar, Kristin Boye, Christopher A. Francis: Diverse and unconventional methanogens, methanotrophs, and methylotrophs in metagenome-assembled genomes from subsurface sediments of the Slate River floodplain, Crested Butte, CO, USA. In: mSystems, Band 9, Nr. 7, Topic: Genomics and Proteomics, 23. Juli 2024, S. e0031424; doi:10.1128/msystems.00314-24, PMC 11264602 (freier Volltext), PMID 38940520, Epub 28. Juni 2024 (englisch).
Wissenschaftliche Bücher
A. L. Reysenbach: Bergey's Manual of Systematic Bacteriology Volume 1: The Archaea and the deeply branching and phototrophic Bacteria. Hrsg.: D. R. Boone, R. W. Castenholz. 2. Auflage. Springer Verlag, New York 2001, ISBN 978-0-387-98771-2, Class IV. Thermoplasmata class. nov., S.169 (englisch).S. 169.
G. M. Garrity, J. G. Holt: Bergey's Manual of Systematic Bacteriology Volume 1: The Archaea and the deeply branching and phototrophic Bacteria. Hrsg.: D. R. Boone, R. W. Castenholz. 2. Auflage. Springer Verlag, New York 2001, ISBN 978-0-387-98771-2, Phylum AII. Euryarchaeota phy. nov., S.169 (englisch).S. 169.
↑ abcChrista Schleper, G. Puehler, I. Holz, A. Gambacorta, D. Janekovic, U. Santarius, H. P. Klenk, Wolfram Zillig: Picrophilus gen. nov., fam. nov.: a Novel Aerobic, Heterotrophic, Thermoacidophilic Genus and Family Comprising Archaea Capable of Growth around pH 0. In: ASM Journals: Journal of Bacteriology, Band 177, Nr. 24, Dezember 1995, S. 7050–7059; doi:10.1128/jb.177.24.7050-7059.1995, PMID 8522509, PMC 177581 (freier Volltext).
↑A. L. Reysenbach: Bergey's Manual of Systematic Bacteriology Volume 1: The Archaea and the deeply branching and phototrophic Bacteria. Hrsg.: D. R. Boone, R. W. Castenholz. 2. Auflage. Springer Verlag, New York 2001, ISBN 978-0-387-98771-2, Class IV. Thermoplasmata class. nov., S. 169, S.169 (englisch).
↑
Eugene Rosenberg, Edward F. DeLong, Stephen Lory, Erko Stackebrandt, Fabiano Thompson (Hrsg.): The Prokaryotes. Other Major Lineages of Bacteria and the Archaea. In: Springer Reference, 4. Auflage, Springer-Verlag Berlin Heidelberg 2014, ISBN 978-3-642-38953-5; doi:10.1007/978-3-642-38954-2 (englisch).
↑
Joel Cracraft, Michael J. Donoghue (Hrsg.): Assembling the tree of life. Oxford University Press US, 2004, ISBN 0-19-517234-5, Online-ISBN 978-0197700235, Print-ISBN 978-0195172348, S.58–59, doi:10.1093/oso/9780195172348.001.0001 (englisch).
↑ abc
Xueju Lin, Kim M. Handley, Jack A. Gilbert, Joel E. Kostka: Metabolic potential of fatty acid oxidation and anaerobic respiration by abundant members of Thaumarchaeota and Thermoplasmata in deep anoxic peat. In: ISME Journal. 9. Jahrgang, Nr.12, 22. Mai 2015, S.2740–2744, doi:10.1038/ismej.2015.77, PMID 26000553, PMC 4817634 (freier Volltext) – (englisch).
↑ abcdefghijklmnopqrstuvwxy
Peng-Fei Zheng, Zhanfei Wei, Yingli Zhou, Qingmei Li, Zhao Qi, Xiaoping Diao, Yong Wang: Genomic Evidence for the Recycling of Complex Organic Carbon by Novel Thermoplasmatota Clades in Deep-Sea Sediments. In: ASM Journals: mSystems, Band 7, Nr. 3, 28. Juni 2022, S. e0007722; doi:10.1128/msystems.00077-22, PMC 9239135 (freier Volltext), PMID 35430893, Epub 18. April 2022 (englisch). Siehe insbes. Tbl. 1.
↑
Anna Shoemaker, Andrew Maritan, Su Cosar, Sylvia Nupp, Ana Menchaca, Thomas Jackson, Aria Dang, Bonnie K. Baxter, Daniel R. Colman, Eric C. Dunham, Eric S. Boyd: Wood-Ljungdahl pathway encoding anaerobes facilitate low-cost primary production in hypersaline sediments at Great Salt Lake, Utah. In: FEMS Microbiology Ecology. 100. Jahrgang, Nr.8, 12. Juli 2024, S.fiae105, doi:10.1093/femsec/fiae105, PMID 39054286, PMC 11287216 (freier Volltext) – (englisch).
↑ abcdef
Spencer Diamond, Adi Lavy, Alexander Crits-Christoph, Paula B. Matheus Carnevali, Allison Sharrar, Kenneth H. Williams, Jillian F. Banfield: Soils and sediments host Thermoplasmata archaea encoding novel copper membrane monooxygenases (CuMMOs). In: ISME Journal. 16. Jahrgang, Nr.5, 5. Januar 2022, S.1348–1362, doi:10.1038/s41396-021-01177-5, PMID 34987183, PMC 9038741 (freier Volltext) – (englisch). Siehe insbes. Fig. 2.
↑
Francesco Candeliere, Laura Sola, Stefano Raimondi, Maddalena Rossi, Alberto Amaretti: Good and bad dispositions between archaea and bacteria in the human gut: New insights from metagenomic survey and co-occurrence analysis. In: Synthetic and Systems Biotechnology. 9. Jahrgang, Nr.1, 3. Januar 2024, S.88–98, doi:10.1016/j.synbio.2023.12.007, PMID 38292760, PMC 10824687 (freier Volltext) – (englisch).
↑ abc
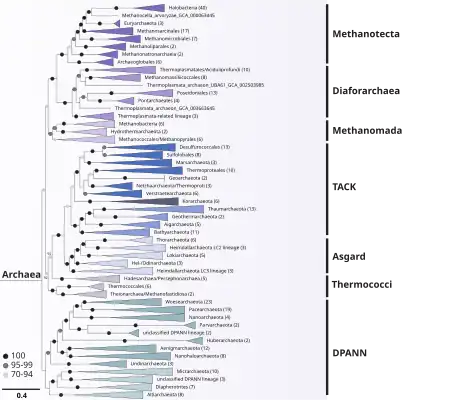
Edmund R. R. Moody, Tara A. Mahendrarajah, Nina Dombrowski, James W. Clark, Celine Petitjean, Pierre Offre, Gergely J. Szöllősi, Anja Spang: An estimate of the deepest branches of the tree of life from ancient vertically evolving genes. In: eLife, Band 11, Nr. e66695, Epub 22. Februar 2022, Stand: 2. März 2022; doi:10.7554/eLife.66695, PMC 8890751 (freier Volltext), PMID 35190025, PDF (englisch). Dazu:
Fig. 4: Phylogenie der Archaea und Bakterien, abgeekeitet aus 27 Markergenen.
Edmund R. R. Moody: Protein evolution and the early history of life. Dissertation, School of Biological Sciences, University of Bristol, Februar 2022, 206 Seiten; PDF (englisch).
Tara A. Mahendrarajah: Tracing the evolutionary history of cellular life. Doktorarbeit, Universität Utrecht, 13. September 2024, 317 Seiten; PDF (englisch).
↑ abcdefghijklmnopqrstuv
Laura A. Zinke, Paul N. Evans, Christian Santos-Medellin, Alena L. Schroeder, Donovan H. Parks, Ruth K. Varner, Virginia I. Rich, Gene W. Tyson, Joanne B. Emerson: Evidence for non‐methanogenic metabolisms in globally distributed archaeal clades basal to the Methanomassiliicoccales. In: Environmental Microbiology, Band 23, Nr. 1, Januar 2021, S. 340–357; doi:10.1111/1462-2920.15316, PMID 33185945, Epub 27. November 2020 (englisch). Anm.: Die GTDB-Ordnung o__RBG-16-68-12 wird teilweise mit RBG-16 abgekürzt (andere GTDB-Ordungen beginnend mit ‚o__RBG-16-…‘ sind nicht Gegenstand dieser Studie.
↑ abcde
Wenzhe Hu, Jie Pan, Bin Wang, Jun Guo, Meng Li, Meiying Xu: Metagenomic insights into the metabolism and evolution of a new Thermoplasmata order (Candidatus Gimiplasmatales). In: Environmental Microbiology; Band 23, Nr. 7, Special Issue on Ecophysiology of Extremophiles, Juli 2021, S. 3695-3709; doi:10.1111/1462-2920.15349, ResearchGate:347764720, Epub 9. Dezember 2020 (englisch). Dazu:
↑
Xiaosi Su, Geng Cui, Huang Wang, Zhenxue Dai, Nam Chil Woo, Wenzhen Yuan: Biogeochemical zonation of sulfur during the discharge of groundwater to lake in desert plateau (Dakebo Lake, NW China). In: Environmental Geochemistry and Health, Band 40, Nr. 3, Juni 2018, S. 1051–1066; doi:10.1007/s10653-017-9975-9, ResearchGate:316957900, PMID 28502020 (englisch).
↑ ab
Xiuran Yin, Guowei Zhou, Mingwei Cai, Qing-Zeng Zhu, Tim Richter-Heitmann, David A. Aromokeye, Yang Liu, Rolf Nimzyk, Qingfei Zheng, Xiaoyu Tang, Marcus Elvert, Meng Li, Michael W. Friedrich: Catabolic protein degradation in marine sediments confined to distinct archaea. In: The ISME Journal, Band 16, Nr. 6, Juni 2022, S. 1617-1626; doi:10.1038/s41396-022-01210-1, PMC 9123169 (freier Volltext), PMID 35220398, Epub 26. Februar 2022 (englisch).
↑
Panagiotis S. Adam, Guillaume Borrel, Céline Brochier-Armanet, Simonetta Gribaldo: The growing tree of Archaea: new perspectives on their diversity, evolution and ecology. In: The ISME Journal, Band 12, Nr. 11, 4. August 2017, S. 2407–2425; doi:10.1038/ismej.2017.122, PMC 5649171 (freier Volltext), PMID 28777382 (englisch).
↑ abcdefghijklm
Yang Yuan, Jun Liu, Tao-Tao Yang, Shao-Ming Gao, Bin Liao, Li-Nan Huang: Genomic Insights into the Ecological Role and Evolution of a Novel Thermoplasmata Order, “Candidatus Sysuiplasmatales”. In: ASM Journals: Applied and Environmental Microbiology, Band 87, Nr. 22, S. e0106521, 28. Oktober 2021; doi:10.1128/AEM.01065-21, PMC 8552897 (freier Volltext), PMID 34524897, Epub 15. September 2021 (englisch).
↑
Heng Zhou, Dahe Zhao, Shengjie Zhang, Qiong Xue, Manqi Zhang, Haiying Yu, Jian Zhou, Ming Li, Sumit Kumar, Hua Xiang: Metagenomic insights into the environmental adaptation and metabolism of Candidatus Haloplasmatales, one archaeal order thriving in saline lakes. In: Environmental Microbiology, Band 24, Nr. 5, Mai 2022, S. 2239-2258; doi:10.1111/1462-2920.15899, PMID 35048500, Epub 20. Januar 2022 (englisch). Dazu:
↑Zur aus TACK und Asgard bestehenden Klade siehe Supergruppe Proteoarchaea. Nach dieser Phylogenie sind die Hadarchaeota und Thermococci keine Mitglieder der Methanobacteriati, sondern stehen nächst den Proteoarchaea.
↑in der NCBI-Taxonomie direkt unter Thermoplasmatota, nicht unter Thermoplasmata
↑
Der vermittels Metagenomik-Daten vorgeschlagene Stamm ARK-15 gehört in der NCBI-Taxonomie zur Gattung Aciduliprofundum (Ordnung Aciduliprofundales/DHVE2). Die GTDB stellt ihn jedoch in eine eigene Ordnung ARK-15 innerhalb der (gemeinsamen) Klasse Thermoplasmata.
↑ abcd
Die ursprünglichen Bezeichnungen Haloplasmatales, Haloplasmataceae und Haloplasma sind wegen Doppelvergabe ungültig und wurden geändert. Haloplasmatales Raineyet al. 2008, Haloplasmataceae Raineyet al. 2008 und HaloplasmaAntuneset al. 2008 bezeichnen bereits eine Bakterienordnung, -familie bzw. -gattung der Bacilli (Firmicutes, nach GTDB) oder Mollicutes (Mycoplasmatota nach LPSN, NCBI).
↑ abcde
Die Ordnungen o__PWKY01 (GTDB) und Ca. Halarchaeoplasmatales (NCBI-Taxonomie), deren Familien f__PWKY01 (GTDB) und Ca. Halarchaeoplasmataceae (NCBI), deren Gattungen PWKY01 (GTDB) und Ca. Halarchaeoplasma (NCBI), inklusive deren Spezies PWKY01 sp007133925 (GTDB) und Ca. Halarchaeoplasma halalkaliphilum (NCBI) sind jeweils synonym aufgrund des gemeinsamen Referenzstamms CSSed165cm_322 dieser Spezies.


{kind=link}