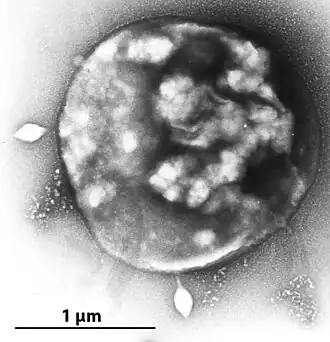
Ein ungefähres Bild der lappigen Form von Archaeen der Gattung Sulfolobus.
Sulfolobaceae ist die Bezeichnung einer FamiliehyperthermophilerArchaea der Ordnung Sulfolobales aus dem PhylumThermoproteota im ReichThermoproteati.[1][2][A. 1]
Einige Vertreter sind darüber hinaus acidophil.
Die Familie wurde (nach einem Vorschlag von Zilliget al. aus dem Jahr 1986) im Jahr 1989 von Karl O. Stetter vom Archaeenzentrum der Universität Regensburg im Bergey’s Manual of Systematic Bacteriology erstbeschrieben.
Die Arten der Typusgattung Sulfolobus wachsen in vulkanischen Quellen, wobei das optimale Wachstum bei einem pH-Wert von 2 bis 3 und Temperaturen von 75 bis 80 °C stattfindet, weshalb sie als acidophil bzw. thermophil gelten. Sulfolobus-Zellen sind unregelmäßig geformt und begeißelt.
Von der Gattung Sulfolobus wurde 2018 von Sakai & Kuroswa die neue Gattung Saccharolobus mit den Spezies S. solfataricus sowie S. shibatae abgespalten; und die Spezies Sulfolobus tokodaii wurde in diesem Jahr von Tsuboi et al. in die bereits bestehende Gattung Sulfurisphaera verschoben.[3] Im Gegensatz dazu wurde ein Vorschlag, auch Sulfolobus islandicus in die Gattung Saccharolobus zu verschieben, im Allgemeinen nicht gefolgt. Alle drei Gattungen, Sulfolobus, Saccharolobus und Sulfurisphaera gehören zur Familie Sulfolobaceae.[4][5][6]
Die Artnamen der Sulfolobaceae wurden und werden in der Regel nach dem Ort benannt, an dem sie zuerst isoliert wurden, z. B. wurde Saccharolobus solfataricus (damals noch Sulfolobus solfataricus genannt) zuerst im Vulkan Solfatara (Pozzuoli bei Neapel, Italien) isoliert. Andere Arten sind weltweit in Gebieten mit vulkanischer oder geothermischer Aktivität zu finden, z. B. in geologischen Formationen, die als Schlammtöpfe oder Solfatare bezeichnet werden.
Ökologie
Die Sulfolobaceae-Arten sind thermophil und kommen oft in heißen Quellen, hydrothermalen Schloten, Schlammtöpfen und vulkanisch aktiven Regionen vor, wobei alleine die Gattung Sulfolobus bzw. die beiden ihr nahestehenden Gattungen Saccharolobus und Sulfurisphaera fast überall vorkommen, wo es vulkanische Aktivitäten gibt.
Die Art Saccharolobus solfataricus wurde etwa u. a. in folgenden vulkanischen Gebieten gefunden: im Yellowstone-Nationalpark, am Mount St. Helens, auf Island, in Italien (Pozzuoli bei Neapel) und Russland (Kamtschatka).
Sulfolobaceae gedeihen in einem Temperaturbereich von etwa 80 °C (allgemein 40 bis 95 °C). Sie sind bei pH-Werten zwischen 1 und 6 zu finden, bestimmte Arten gelten daher als acidophil (säureliebend). Sulfurisphaera tokodaii, wurde in einem sauren Thermalbad der Beppu Hot Springs, Präfektur Ōita, Kyushu, Japan, gefunden. Manche Arten, wie Saccharolobus solfataricus, sind mit Schwefelvorkommen assoziiert. Andere, wie Metallosphaera prunae, wurden auf schwelendem Abfallmaterial aus Bergwerken gefunden, wo sie einen lithoautotrophen Stoffwechsel betreiben.[7]
Sedimente aus ~90 m Tiefe unter dem Meeresboden am peruanischen Kontinentalrand werden von intakten archaeenspezifischen Tetraethern[8] dominiert, die auf Sulfolobaceae hindeuten (s. u.). Offenbar besteht ein bedeutender Teil mikrobiellen Gemeinschaft im Sediment aus Archaeen, die taxonomisch mit der Ordnung Sulfolobales verbunden sind.[9]
Sulfolobaceae sind auch an der Biofilmbildung beteiligt. Die Analyse von Biofilmen, die von verschiedenen Arten produziert werden, hat gezeigt, dass diese Arten nur sehr wenige regulierende Proteine gemeinsam haben. Man vermutet daher, dass es möglicherweise mehrere unterschiedliche Regulierungsmechanismen für die Biofilmbildung gibt. Die Bildung von turmartigen Biofilmstrukturen werden bei Sulfolobus acidocaldarius beobachtet, während Saccharolobus solfataricus teppichartige Biofilmen ausbildet.[10]
Stoffwechsel
Sulfolobaceae-Arten weisen eine breite Palette von Stoffwechselarten auf, darunter aerobe, fakultativ anaerobe oder obligat anaerobe. Diese gehen einher mit einer chemoheterotrophen, lithoautotrophen oder mixotrophen Lebensweise.
Einige Arten weisen eine metabolische Flexibilität auf, indem sie je nach den verfügbaren Energiequellen mehrere verschiedene Stoffwechselwege nutzen können, während andere nur eine begrenzte Auswahl an metabolischen Optionen haben.[11]
Chemoheterotrophe Sulfolobaceae gewinnen ihre Energie, indem sie einfache reduzierte Kohlenstoffverbindungen oxidieren, was mit Hilfe oxidierter Formen von Schwefel geschieht. Zu den fraglichen Kohlenstoffverbindungen gehören u. a. D-Glukose, D-Galaktose und andere gängige Zuckerarten, aber auch Aminosäuren und andere komplexe Moleküle.
Beim lithoautotrophen Stoffwechsel hingegen wird die Energie aus der Oxidation von reduzierten Verbindungen wie elementarem Schwefel, Schwefelerzen und anderen reduzierten Schwefelverbindungen oder molekularem Wasserstoff gewonnen.
Heterotrophes Wachstum wurde jedoch nur in Gegenwart von Sauerstoff beobachtet. Die wichtigsten Stoffwechselwege sind ein glykolytischer Weg, ein Pentosephosphatweg und der TCA-Zyklus (Citratzyklus).
Sulfolobaceae bevorzugen mit Ausnahme der Gattung Acidianus Umgebungen mit niedrigem Natriumchloridgehalt. Glykogen wird als langfristiger Kohlenstoff- und Energiespeicher verwendet.
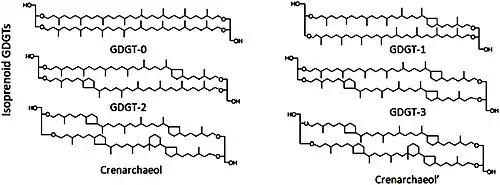
Molekulare Strukturen von isoprenoiden Glycerindialkylglycerin-Tetraethern (GDGTs) mit 0 bis 4 Cyclopentan-Ringen: GDGT-0 alias Caldarchaeol bis GDGT-4 und Crenarchaeole
Alle Archaeen haben Lipide mit Etherbindungen zwischen der Kopfgruppe und den Seitenketten, die im Vergleich zu den bakteriellen und eukaryotischen estergebundenen Lipiden hitze- und säurebeständiger sind.
Unter den Archaeen haben die Sulfolobales ansonsten ungewöhnliche Tetraetherlipide.[8] Ihre etherverknüpften Lipide sind kovalent über die „Doppelschicht“ (Bilayer) hinweg verbunden und bilden so Tetraether – technisch gesehen bilden die Tetraether also eine Monoschicht und keine Doppelschicht. Die Tetraether tragen dazu bei, dass diese Sulfolobaceae-Arten sowohl extreme Säure als auch hohe Temperaturen überleben.[12]
Morphologie
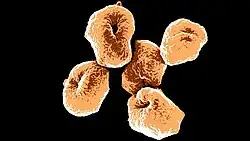
Die Zellformen der Sulfolobaceae-Arten ist entweder regelmäßig, unregelmäßig oder gelappte Kokkenförmig. Ihre Größe liegt zwischen 0,5 und 2 μm im Durchmesser. Es wurden mehrere verschiedene Oberflächenanhänge beobachtet, darunter bei beweglichen (motilen) Arten Archaellen, die typischerweise bei Nahrungsmittelknappheit (Hunger) auftreten.[13]
Einzigartig bei Sulfolobus acidocaldarius sind die archaeenspezifischen adhäsivenPili, die für die Oberflächenanhaftung bei der Biofilmbildung wichtig sind.[14]
Bevor Filamente dieser Archaeen-Oberflächenanhängsel aufgebaut werden können erfolgt die Expression von Typ-IV-Pilin durch eine spezielle Typ-IV-Präpilin-Signalpeptidase. In Saccharolobus solfataricus wurde eine zuckerbindende Oberflächenstruktur gefunden, die als „Bindosom“ bezeichnet wird. Wenn solch ein spezielles Bindosomensystem zusammengebaut wird, sind sie am Zuckertransport aktiv beteiligt.[15]
Bestimmte Stämme von Sulfolobus islandicus bilden proteinhaltige Toxine, die so genannten Sulfolobicine.
Diese Toxine können einen Wettbewerbsvorteil darstellen, da sie das Wachstum von nicht toxinbildenden Stämmen von S. islandicus und bestimmten anderen Sulfolobaceae-Arten hemmen. Andererseits werden andere Arten, wie z. B. Sulfolobus acidocaldarius nicht gehemmt. Das Gen, das für Sulfolobicine kodiert, wurde auch bei anderen Sulfolobaceae-Arten identifiziert.[16]
Modellorganismen
Sulfolobus und verwandte Arten der Familie Sulfolobaceae dienen als Modell für die Untersuchung der molekularen Mechanismen der DNA-Replikation – auch bei Eukaryoten.
Als 1996 das erste Archaeen-Genom, Methanococcus jannaschii (Methanobacteriota, früher Euryarchaeota), vollständig sequenziert wurde, stellte man fest, dass die an der DNA-Replikation, Transkription und Translation beteiligten Gene im Genom dieser Art mehr mit ihren Gegenstücken in Eukaryoten als mit denen in anderen Prokaryoten verwandt waren.
Im Jahr 2001 wurde die erste Genomsequenz, Saccharolobus solfataricus (damals Sulfolobus solfataricus genannt), Stamm P2, veröffentlicht.
Im Genom des Stamms P2 wurden dabei ebenfalls Gene gefunden, die mit der Chromosomenreplikation zusammenhängen und die mehr mit denen von Eukaryonten verwandt sind. Unter diesen befinden sich Gene, die eine DNA-Polymerase, Primase (einschließlich zweier Untereinheiten), MCM, CDC6/ORC1 (Helikasen), CDC6/ORC1 (origin recognition complex, siehe Replikation §Eukaryotische Replikation), RPA & RPC (Replication protein A bzw. C) und PCNA.
Im Jahr 2004 wurden die Ursprünge der DNA-Replikation von Saccharolobus solfataricus und Sulfolobus acidocaldarius identifiziert.
Es zeigte sich, dass beide Arten (je) zwei Replikationsursprünge in ihrem Genom haben. Dies war das erste Mal, dass in einer prokaryontischen Zelle mehr als ein einziger DNA-Replikationsursprung nachgewiesen werden konnte.
Der Mechanismus der DNA-Replikation in Archaeen ist evolutionär konserviert und ähnelt dem von Eukaryoten. Sulfolobus und Saccharolobus werden nun als Modell verwendet, um die molekularen Mechanismen der DNA-Replikation in Archaeen zu untersuchen.
Weil das System der DNA-Replikation in Archaeen viel einfacher ist als in Eukaryoten, wurde vorgeschlagen, dass Archaeen als Modell für die Untersuchung der viel komplexeren DNA-Replikation in Eukaryoten verwendet werden könnten.
Genom-Sequenzierungen
Unter anderem wurden bisher (Stand 2005) die vollständigen Genome folgender Arten sequenziert:
Sulfolobus acidocaldarius, Stamm DSM 639 ;(2.225.959 bp, d. h. Basenpaare),[17]
Saccharolobus solfataricus hat ein zirkuläres Chromosom mit 2.992.245 Basenpaaren.
Die weitere sequenzierte Art, Sulfurisphaera tokodaii, hat ebenfalls ein zirkuläres Chromosom, das mit 2.694.756 bp jedoch etwas kleiner ist.
Beiden Arten fehlen die Gene ftsZ[20] und minD,[21] was auch für sequenzierte Thermoproteota (früher „Crenarchaeota“) charakteristisch ist.
Sie kodieren auch für die Citrat-Synthase und zwei Untereinheiten der 2-Oxosäure:Ferredoxin-Oxidoreduktase,[22] die die gleiche Rolle wie die α-Ketoglutarat-Dehydrogenase im TCA-Zyklus (alias Tricarbonsäurezyklus, Krebs-Zyklus oder Citratzyklus) spielt. Dies deutet darauf hin, dass diese Spezies über ein TCA-Zyklus-System verfügt, das dem der Mitochondrien von Eukaryonten ähnelt.
Andere Gene in der Atmungskette, die an der Produktion von ATP beteiligt sind, ähneln jedoch nicht denen von Eukaryonten. Ein solches Beispiel ist Cytochrom c, das bei Eukaryonten eine wichtige Rolle bei der Elektronenübertragung auf Sauerstoff spielt. Dies wurde auch bei Aeropyrum pernix (Thermoprotei), Stamm K1, festgestellt. Da dieser Schritt für einen aeroben Mikroorganismus der Sulfolobaceae wichtig ist, verwendet er wahrscheinlich ein anderes Molekül für dieselbe Funktion oder hat einen anderen Stoffwechselweg.
Reaktion auf DNA-Schäden
Die Exposition von Saccharolobus solfataricus oder Sulfolobus acidocaldarius gegenüber den DNA-schädigenden Agenzien wie UV-Strahlung, Bleomycin oder Mitomycin C induziert eine zelluläre Aggregation;[23][24] andere physikalische Stressfaktoren wie pH- oder Temperaturverschiebung lösten jedoch keine Aggregation aus.
Dies deutet darauf hin, dass die Herbeiführung (Induktion) einer Aggregation spezifisch durch DNA-Schäden verursacht wird.[24]
Ajon et al. zeigten 2011, dass die UV-induzierte zelluläre Aggregation bei S. acidocaldarius den Austausch chromosomaler Marker mit hoher Frequenz vermittelt.
Die Rekombinationsraten übertrafen die von normalen, „nicht induzierten“ Kulturen um bis zu drei Größenordnungen.[23]
Wood et al. zeigten 1997 ebenfalls, dass UV-Bestrahlung die Häufigkeit von Rekombinationen aufgrund von genetischem Austausch in Sulfolobus acidocaldarius erhöht.[25]
Fröls et al. sowie Ajon et al. stellten 2008–2011 die Hypothese auf, dass der durch UV-Strahlung induzierte DNA-Transferprozess und die anschließende homologe Rekombinationsreparatur einen wichtigen Mechanismus zur Aufrechterhaltung der Chromosomenintegrität in Sulfolobus acidocaldarius und Saccharolobus solfataricus darstellen.[24][26][23]
Diese Reaktion kann eine primitive Form der sexuellen Interaktion angesehen werden, ähnlich wie die besser untersuchte bakterielle Transformation, die ebenfalls mit einem DNA-Transfer zwischen Zellen verbunden ist und zu einer homologen rekombinativen Reparatur von DNA-Schäden führt.[27][28]
Das ups Operon
Das ups-Operon von acidocaldarius acidocaldarius wird durch UV-Bestrahlung stark induziert.[29][30] Die von diesem Operon kodiertenPili werden zur Förderung der Zellaggregation eingesetzt, die für den anschließenden DNA-Austausch zwischen den Zellen notwendig ist, was zu homologer Rekombination führt. Eine Studie des ups-Operons von acidocaldarius acidocaldarius durch Marleen van Wolferen et al. (2015) zeigte, dass eines der Gene des Operons, saci-1497, für eine Endonuklease III kodiert, die UV-geschädigte DNA kappt. Ein anderes Gen des Operons, saci-1500, kodiert für eine RecQ-ähnliche Helikase,[31] die in der Lage ist, homologe Rekombinations-Zwischenprodukte wie Holliday Junctions abzuwickeln.
Das Team schlug daher vor, dass Saci-1497 und Saci-1500 in einem auf homologer Rekombination basierenden DNA-Reparaturmechanismus fungieren, der übertragene DNA als Vorlage verwendet. Man nimmt also an, dass das ups-System in Kombination mit homologer Rekombination eine DNA-Schadensreaktion darstellt, die Sulfolobales vor DNA-schädigenden Bedrohungen rettet.[30]
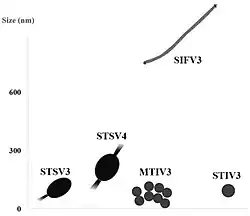
Morphologie und Größe der in den heißen Quellen von Tengchong-Gebiet gefundenen Sulfolobus- und Metallosphaera-Viren (SIFV3 = SSRV2).
Viren können die extrem sauren und heißen Bedingungen, in denen Sulfolobaceae leben, nicht überstehen. Sie nutzen die Mitglieder dieser Familie daher nicht nur, um sich zu vermehren. Sie nutzen die für sie viel verträglicheren Bedingungen im Innern ihrer Wirtszellen, um überhaupt zu überdauern.
Da der Wirt ihnen Schutz bietet, ist es für die Viren nicht vorteilhaft, die Wirte zu zerstören (lysieren), wenn sie dadurch den reuen Umweltbedingungen ausgesetzt werden.
Die Viren der Sulfolobaceae (wie etwa von der Familie Fuselloviridae) sind daher vorzugsweise gemäßigt oder permanent lysogen. Permanente Lysogene unterscheiden sich von anderen lysogenen Viren dadurch, dass die Wirtszellen nach der Produktion ihrer Nachkommen nicht lysieren in den lysogenen Zustand zurückkehren.
Da die Archaeen wie Sulfolobus infizierenden Viren eine Strategie anwenden müssen, um einer längeren direkten Exposition gegenüber den Umgebungen, in denen ihre Wirte leben, zu entgehen.
Dies könnte einige ihrer einzigartigen Eigenschaften erklären.
Fuselloviridae sind u. a. darin einzigartig, dass ihre Gene für die Strukturproteine ständig transkribiert werden.
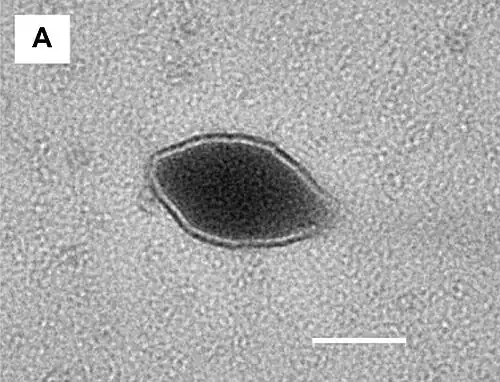
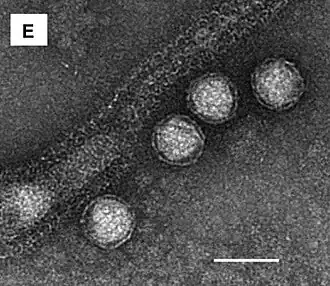
Die Sulfolobaceae werden von einer Reihe sehr vielgestaltiger Viren parasitiert. Die folgenden Bilder stellen eine Reihe von Viren vor, die in den sauer-heißen Quellen im Tengchong-Gebiet, Yunnan (China) gefunden wurden.[46]
Zitronen-, spindel-, tropfen- und flaschenförmige Archaeenviren
„Sulfolobus virus STSV3“ („Sulfolobus tailed spindle-shaped virus 3“), nicht klassifiziert
„Sulfolobus virus STSV4“ („Sulfolobus tailed spindle-shaped virus 4“), nicht klassifiziert
Infektionen mit spindelförmigen Sulfolobaceae-Viren (SSV). REM-Aufnahme von S. solfataricus Stamm Gθ, nicht infiziert (A) bzw. mit SSVRH infiziert (B); (C)TEM-Aufnahme von SSVRH-Virionen, (D) TEM von SSVRH-Virionen mit infiziertem S. solfataricus Stamm Gθ (unten links)[48]
Halo-Bildung bei S. solfataricus Stamm Gθ für verschiedene SSVs. Die Virusanfälligkeit wird durch die Bildung von Halos der Wachstumshemmung um einen Spot des Viruskonzentrats bestimmt, das auf eine Petrischale mit S. solfataricus Gθ aufgetragen wurde. Die Halo-Formen sind typisch für die einzelnen SSV-Vertreter.[48]
Auch S. ohwakuensis ist für SSV-Infektion empfänglich. SSV3, SSVRH und SSVK1 erzeugen Halos auf S. ohwakuensis, SSV1, SSV2 und SSVL1 hemmen deren Wachstum nicht.[48]
Repräsentative Virusanfälligkeit der drei Wirtsklassen von Sulfolobaceae. Die Virusinfektion wurde wie in Abbildung 2 bestimmt. (A) Vollständig anfälliger Wirt: Alle 6 getesteten SSV hemmen das Wachstum eines von Adam J. Clore aus dem Lassen-Volcanic-Nationalpark isolierten Sulfolobaceae-Stammes. (B) Vollständig resistenter Wirt: Keine SSV hemmen das Wachstum von Sulfurisphaera tokodaii [Sulfolobus tokodaii]. (C) Selektiv anfälliger Wirt: 4 von 6 SSV-Vertretern hemmen das Wachstum von „Sulfolobus islandicus“ Stamm HVE 10/4. Die Flecken auf den Platten sind (im Uhrzeigersinn vom Pfeil aus): Triton X-100 (positive Kontrolle), SSV1, SSV2, SSV3, SSVL1, SSVK1, SSVRH, und steriles H2O (negative Kontrolle).[48]
Wegen der Aufspaltung der Gattung Sulfolobus tragen viele Viren diesen Gattungsnamen in ihrer Bezeichnung, auch wenn sie (zuerst) in Wirten einer abgespaltenen Gattung nachgewiesen wurden.
↑Die Genome Taxonomy Database (GTDB) verweist die Ordnung Sulfolobales nicht in die Klasse Thermoprotei, sondern in eine eigene Klasse Thermoprotei_A (manchmal auch als Sulfolobia bezeichnet).
↑Die LPSN sieht Sulfosphaerellus zwar als eigene monotypische Gattung in Sulfolobaceae ohne Familienzuordnung, NCBI-Taxonomie betrachtet Sulfosphaerellus thermoacidophilum als gemäß He et al. (2004) ein Synonym von Acidianus tengchongensis.
↑In der GTDB sind HO1-1 und JCM 8857 verschiedene Stämme der Spezies Metallosphaera hakonensis
↑Nach der GTDB gehört der Stamm UBA165 ebenfalls zur Spezies Metallosphaera sedula
↑In der GTDB gehört der Stamm AS-7 zur Spezies Metallosphaera javensisHofmannet al. 2022, es gibt dort keinen Unterschied. Die LPSN listet den Stamm AS-7 nicht, ist also neutral.
↑In der GTDB sind IC-006 und JCM 16833 verschiedene Stämme der Spezies Sulfuracidifex tepidarius
↑In der GTDB sind 7 und NBRC 100140 verschiedene Stämme der Spezies Sulfurisphaera tokodaii
↑Die Kandidatengattung „Ca. Aramenus“ wird in der GTDB und der NCBI-Taxonomie der Familie Sulfolobaceae zugeordnet, in der LPSN wird sie als Sulfolobales incertae sedis gesehen.
↑Nach der GTDB gehört der Stamm AZ2 ebenfalls zur Spezies „Ca. Aramenus sulfurataquae“.
↑Die GTDB spaltet den Stamm AZ1 in den eigentlichen Referenzstamm AZ1-illumnia und einen zweiten Stamm AZ1-454 zur selben Spezies.
↑auch als „Sulfolobus newzealandicus droplet-shaped virus“ verschrieben; das Virus und seine Spezies wurden inzwischen vom ICTV deklassifiziert, da (noch) keine vollständige Genomsequenz vorliegt.
Kinshuk Chandra Nayak: Comparative genome sequence analysis of Sulfolobus acidocaldarius and 9 other isolates of its genus for factors influencing codon and amino acid usage. In: Gene, band 513, Nr. 1, 15. Januar 2013, S. 163-173; doi:10.1016/j.gene.2012.10.024, PMID 23124074. Epub 30. Oktober 2012 (englisch).
Andrés Santos, Pablo Bruna, Jaime Martinez-Urtaza, Francisco Solis, Bernardita Valenzuela, Pedro Zamorano, Leticia Barrientos: Two Archaeal Metagenome-Assembled Genomes from El Tatio Provide New Insights into the Crenarchaeota Phylum. In: MDPI: Genes, Band 12, Nr. 3, 9. März 2021, S. 391; doi:10.3390/genes12030391 (englisch).
Quellen
Benutzte Literatur
Judicial Commission of the International Committee on Systematics of Prokaryotes: The nomenclatural types of the orders Acholeplasmatales, Halanaerobiales, Halobacteriales, Methanobacteriales, Methanococcales, Methanomicrobiales, Planctomycetales, Prochlorales, Sulfolobales, Thermococcales, Thermoproteales and Verrucomicrobiales are the genera Acholeplasma, Halanaerobium, Halobacterium, Methanobacterium, Methanococcus, Methanomicrobium, Planctomyces, Prochloron, Sulfolobus, Thermococcus, Thermoproteus and Verrucomicrobium, respectively. Opinion 79. In: International Journal of Systematic and Evolutionary Microbiology, Ban 55, Nr. 1, 1. Januar 2005, S. 517–518; doi:10.1099/ijs.0.63548-0, PMID 15653928 (englisch).
Thomas D. Brock, Katherine M. Brock, Robert T. Belly, Richard L. Wei ss: Sulfolobus: a new genus of sulfur-oxidizing bacteria living at low pH and high temperature. In: Archiv für Mikrobiologie, Band 84, Nr. 1, März 1972, S. 54–68; doi:10.1007/BF00408082, PMID 4559703 (englisch).
Karl O. Stetter: Bergey's Manual of Systematic Bacteriology. Hrsg.: J. T. Staley, M. P. Bryant, N. Pfennig, J. G. Holt. 1. Auflage. Band3. The Williams & Wilkins Co., Baltimore 1989, Order III. Sulfolobales ord. nov. Family Sulfolobaceae fam. nov., S.169 (englisch).
M. Madigan, J. Martinko (Hrsg.): Brock Biology of Microorganisms. 11. Auflage. Prentice Hall, 2005, ISBN 978-0-13-144329-7 (englisch).
↑
Aidan C. Parte, Joaquim Sardà Carbasse, Jan P. Meier-Kolthoff, Lorenz C. Reimer, Markus Göker: List of Prokaryotic names with Standing in Nomenclature (LPSN) moves to the DSMZ. In: International Journal of Systematic and Evolutionary Microbiology. 70. Jahrgang, Nr.11, 2020, ISSN1466-5034, S.5607–5612, doi:10.1099/ijsem.0.004332, PMID 32701423, PMC 7723251 (freier Volltext) – (englisch).
↑ abcd
Hiroyuki D. Sakai, Norio Kurosawa: Saccharolobus caldissimus gen. nov., sp. nov., a facultatively anaerobic iron-reducing hyperthermophilic archaeon isolated from an acidic terrestrial hot spring, and reclassification of Sulfolobus solfataricus as Saccharolobus solfataricus comb. nov. and Sulfolobus shibatae as Saccharolobus shibatae comb. nov. In: International Journal of Systematic and Evolutionary Microbiology, Band 68, Nr. 4, April 2018, S. 1271–1278; doi:10.1099/ijsem.0.002665, PMID 29485400, Epub 27. Februar 2018 (englisch).
↑ ab
Tanja Fuchs, Harald Huber, Kristina Teiner, Siegfried Burggraf, Karl O. Stetter. Metallosphaera prunae, sp. nov., a Novel Metal-mobilizing, Thermoacidophilic Archaeum, Isolated from a Uranium Mine in Germany. In: Systematic and Applied Microbiology, Band 18, Nr. 4, 1995, S. 560-566; doi:10.1016/S0723-2020(11)80416-9 (englisch).
↑ ab
Geraldy Lie Stefanus Liman, Andy A. Garcia, Kristin A. Fluke, Hayden R. Anderson, Sarah C. Davidson, Paula V. Welander, Thomas J. Santangelo: Tetraether archaeal lipids promote long-term survival in extreme conditions. In: Molecular Microbiology, Band 121, Nr. 5, Mai 2024,S. 882-894; doi:10.1111/mmi.15240, PMC 11096074 (freier Volltext), PMID 38372181, Epub 19. Februar 2024 (englisch).
↑
Helen F. Sturt, Roger E. Summons, Kristin Smith, Marcus Elvert, Kai-Uwe Hinrichs: Intact polar membrane lipids in prokaryotes and sediments deciphered by high‐performance liquid chromatography/electrospray ionization multistage mass spectrometry—New biomarkers for biogeochemistry and microbial ecology. In: Rapid Communications in Mass Spectrometry, Band 18, Nr. 6, 2004, S. 617–628; doi:10.1002/(ISSN)1097-0231, PMID 15052572 (englisch).
↑
Andrea Koerdt, Alvaro Orell, Trong Khoa Pham, Joy Mukherjee, Alexander Wlodkowski. Esther Karunakaran, Catherine A. Biggs, Phillip C. Wright, Sonja-Verena Albers: Macromolecular Fingerprinting of Sulfolobus Species in Biofilm: A Transcriptomic and Proteomic Approach Combined with Spectroscopic Analysis. In: Journal of Proteome Research, Band 10, Nr. 9, 2. September 2011, S. 4105-4119; doi:10.1021/pr2003006, PMC 3166137 (freier Volltext), PMID 21761944, Epub 18. Juli 2011 (englisch).
↑
Sonja-Verena Albers, Bettina Siebers: The Family Sulfolobaceae. In: E. Rosenberg et al. (Hrsg.): The Prokaryotes. 4. Auflage, Springer-Verlag, Berlin, Heidelberg, Juli 2014, S. 323-346; doi:10.1007/978-3-642-38954-2_329, ResearchGate:285962786 (englisch).
↑
Zalán Szabó, Musa Sani, Maarten Groeneveld, Benham Zolghadr, James Schelert, Sonja-Verena Albers, Paul Blum, Egbert J. Boekema, Arnold J. M. Driessen: Flagellar Motility and Structure in the Hyperthermoacidophilic Archaeon Sulfolobus solfataricus. In: Journal of Bacteriology, Band 189, Nr. 11, April 2007, S. 4305-4309; doi:10.1128/JB.00042-07, PMC 1913377 (freier Volltext), PMID 17416662 (englisch).
↑
Anna-Lena Henche, Abhrajyoti Ghosh, Xiong Yu, Torsten Jeske, Edward Egelman, Sonja-Verena Albers: Structure and function of the adhesive type IV pilus of Sulfolobus acidocaldarius. In: Environmental Microbiology, Band 14, Nr. 12, Dezember 2012, S. 3188-3202; doi:10.1111/j.1462-2920.2012.02898.x, PMC 3977132 (freier Volltext), PMID 23078543, Epub 19. Oktober 2012 (englisch).
↑
Behnam Zolghadr, Stefan Weber, Zalán Szabó, Arnold J. M. Driessen, Sonja-Verena Albers: Identification of a system required for the functional surface localization of sugar binding proteins with class III signal peptides in Sulfolobus solfataricus. In: Molecular Microbiology, Band 64, Nr. 3, S. 795-806; doi:10.1111/j.1365-2958.2007.05697.x, PMID 17462024 (englisch).
↑
David Prangishvili, Ingelore Holz, Evelyn Stieger, Stephan Nickell, Jakob K. Kristjansson, Wolfram Zillig: Sulfolobicins, Specific Proteinaceous Toxins Produced by Strains of the Extremely Thermophilic Archaeal Genus Sulfolobus. In: Journal of Bacteriology, Band 182, Nr. 10, Mai 2000, S. 2985-2988; doi:10.1128/JB.182.10.2985-2988.2000, PMC 102014 (freier Volltext), PMID 10781574 (englisch).
↑
Lanming Chen, Kim Brügger, Marie Skovgaard, Peter Redder, Qunxin She, Elfar Torarinsson, Bo Greve, Mariana Awayez, Arne Zibat, Hans-Peter Klenk, Roger A. Garrett: The genome of Sulfolobus acidocaldarius, a model organism of the Crenarchaeota. In: Journal of Bacteriology, Band 187, Nr. 14, Juli 2005, S. 4992–4999; doi:10.1128/JB.187.14.4992-4999.2005, PMC 1169522 (freier Volltext), PMID 15995215 (englisch).
↑
Qunxin She, Rama K. Singh, Fabrice Confalonieri, Yvan Zivanovic, Ghislaine Allard, Mariana J. Awayez, Christina C.-Y. Chan-Weiher, Ib Groth Clausen, Bruce A. Curtis, Anick De Moors, Gael Erauso, Cynthia Fletcher, Paul M. K. Gordon, Ineke Heikamp-de Jong, Alex C. Jeffries, Catherine J. Kozera, Nadine Medina, Xu Peng, Hoa Phan Thi-Ngoc, Peter Redder, Margaret E. Schenk, Cynthia Theriault, Niels Tolstrup, Robert L. Charlebois, W. Ford Doolittle, Michel Duguet, Terry Gaasterland, Roger A. Garrett, Mark A. Ragan, Christoph W. Sensen, and John Van der Oost: The complete genome of the crenarchaeon Sulfolobus solfataricus P2. In: PNAS. 98. Jahrgang, Nr.14, 3. Juli 2001, S.7835–7840, doi:10.1073/pnas.141222098, PMID 11427726, PMC 35428 (freier Volltext), bibcode:2001PNAS...98.7835S (englisch).
↑
Piet A. de Boer, Robin E. Crossley, Lawrence I. Rothfield: The essential bacterial cell-division protein FtsZ is a GTPase. In: Nature, band 359, 17. September 1992, S. 254–256; doi:10.1038/359254a0 (englisch).
↑
Piet A. de Boer, Robin E. Crossley, A. R. Hand, Lawrence I. Rothfield: The MinD protein is a membrane ATPase required for the correct placement of the Escherichia coli division site. In: EMBO Journal, Band 10, Nr. 13, Dezember 1991, S. 4371-4280; doi:10.1002/j.1460-2075.1991.tb05015.x, PMC 453190 (freier Volltext), PMID 1836760 (englisch).
↑
Marcus I. Gibson, Percival Yang-Ting Chen, Catherine L. Drennan: A Structural Phylogeny for Understanding 2-Oxoacid Oxidoreductase Function. In: Current Opinion in Structural Biology, Band 41, Dezember 2016, S. 54-61; doi:10.1016/j.sbi.2016.05.011, PMC 5381805 (freier Volltext), PMID 27315560 (englisch).
↑ abc
Małgorzata Ajon, Sabrina Fröls, Marleen van Wolferen, Kilian Stoecker, Daniela Teichmann, Arnold J. M. Driessen, Dennis W. Grogan, Sonja-Verena Albers, Christa Schleper: UV-inducible DNA exchange in hyperthermophilic archaea mediated by type IV pili. In: Molecular Microbiology. 82. Jahrgang, Nr.4, November 2011, S.807–817, doi:10.1111/j.1365-2958.2011.07861.x, PMID 21999488 (englisch).
↑ abc
Sabrina Fröls, Malgorzata Ajon, Michaela Wagner, Daniela Teichmann, Behnam Zolghadr, Mihaela Folea, Egbert J. Boekema, Arnold J. M. Driessen, Christa Schleper, Sonja-Verena Albers: UV-inducible cellular aggregation of the hyperthermophilic archaeon Sulfolobus solfataricus is mediated by pili formation. In: Molecular Microbiology. Band 70, Nr. 4, November 2008, S. 938–952, doi:10.1111/j.1365-2958.2008.06459.x, PMID 18990182 (englisch).
↑
Eric R. Wood, Farnaz Ghané, Dennis W. Grogan: Genetic responses of the thermophilic archaeon Sulfolobus acidocaldarius to short-wavelength UV light. In: Journal of Bacteriology, Band 179, Nr. 18, September 1997, S. 5693–5698; doi:10.1128/jb.179.18.5693-5698.1997, PMC 179455 (freier Volltext), PMID 9294423 (englisch).
↑
Sabrina Fröls, Malcolm F. White, Christa Schleper: Reactions to UV damage in the model archaeon Sulfolobus solfataricus. In: Biochemical Society Transactions, Band 37, Nr. 1, Februar 2009, S. 36–41, doi:10.1042/BST0370036, PMID 19143598 (englisch).
↑
Jeferson Gross, Debashish Bhattacharya: Uniting sex and eukaryote origins in an emerging oxygenic world. In: Biology Direct. 5. Jahrgang, 2010, S.53, doi:10.1186/1745-6150-5-53, PMID 20731852, PMC 2933680 (freier Volltext) – (englisch).
↑
Harris Bernstein, Carol Bernstein: Evolutionary Origin of Recombination during Meiosis. In: BioScience. 60. Jahrgang, Nr.7, 2010, S.498–505, doi:10.1525/bio.2010.60.7.5 (englisch).
↑
Marleen van Wolferen, Małgorzata Ajon, Arnold J. M. Driessen, Sonja-Verena Albers: Molecular analysis of the UV-inducible pili operon from Sulfolobus acidocaldarius. In: MicrobiologyOpen, Band 2, Nr. 2, Dezember 2013, S. 928-937; doi:10.1002/mbo3.128, PMC 3892339 (freier Volltext), PMID 24106028, ResearchGate:257535309 (englisch).
↑ ab
Marleen van Wolferen, Xiaoqing Ma, Sonja-Verena Alber: DNA Processing Proteins Involved in the UV-Induced Stress Response of Sulfolobales. In: Journal of Bacteriology. 197. Jahrgang, Nr.18, 19. August 2015, S.2941–2951, doi:10.1128/JB.00344-15, PMID 26148716, PMC 4542170 (freier Volltext) – (englisch).
↑
H. Zhong, X. Chen, Y. Li, W. Cai: A new genus of thermo- and acidophilic bacteria Sulfosphaerellus (一个嗜热嗜酸细菌的新属硫球菌属Yīgè shì rè shì suān xìjūnde xīn shǔ - liú qiújūn shǔ). In: Acta Microbiologica Sinica, Band 22, 1982, S. 1-7 (chinesisch).
↑
Z. G. He, H. Zhong, Y. Li: Acidianus tengchongensis sp. nov., a new species of acidothermophilic archaeon isolated from an acidothermal spring. In. Current Microbiology, Band 48, 2004, S. 159-163 (englisch).
↑ abcWolfram Zillig, Karl O. Stetter, Simon Wunderl, Wolfgang Schulz, Harro Priess, Ingrid Scholz: The Sulfolobus-“Caldariella” group: Taxonomy on the basis of the structure of DNA-dependent RNA polymerases. In: Archives of Microbiology, Band 125, April 1980, S. 259–269; doi:10.1007/BF00446886 (englisch).
↑
Tang-Jian Peng, Liu Lijun, Chang Liu, Zhi-Fang Yan, Shuang-Jiang Liu, Cheng-Ying Jiang: Metallosphaera tengchongensis sp. Nov., An acidothermophilic archaeon isolated from a hot spring. In: International Journal of Systematic and Evolutionary Microbiology, Band 65, Nr. 2, 1. Februar 2015; doi:10.1099/ijs.0.070870-0, PMID 25404480, ResearchGate:[hzttps://www.researchgate.net/publication/268793692 268793692], Epub November 2014 (englisch).
↑ ab
Xin Dai, Haina Wang, Zhenfeng Zhang, Kuan Li, Xiaoling Zhang, Marielos Mora-López, Chengying Jiang, Chang Liu, Li Wang, Yaxin Zhu, Walter Hernández-Ascencio, Zhiyang Dong, Li Huang: Genome Sequencing of Sulfolobus sp. A20 from Costa Rica and Comparative Analyses of the Putative Pathways of Carbon, Nitrogen, and Sulfur Metabolism in Various Sulfolobus Strains. In: Frontiers in Microbiology, Sec. Extreme Microbiology, Band 7, 30. November 2016; doi:10.3389/fmicb.2016.01902 (englisch).
↑Thomas D. Brock, Katherine M. Brock, Robert T. Belly, Richard L. Weiss: Sulfolobus: A new genus of sulfur-oxidizing bacteria living at low pH and high temperature. In: Archiv für Mikrobiologie. Band 84, März 1972, S. 54–68; doi:10.1007/BF00408082, PMID 4559703 (englisch).
↑
Gertrud Huber, Karl O. Stetter: Sulfolobus metallicus, sp. nov., a Novel Strictly Chemolithoautotrophic Thermophilic Archaeal Species of Metal-Mobilizers. In: Systematic and Applied Microbiology, Band 14, Nr. 4, Oktober 1991, S. 372-378; doi:10.1016/s0723-2020(11)80312-7 (englisch).
↑In der GTDB sind TA-1 und DSM 12421 verschiedene Stämme
↑ ab
Grigory I. Karavaĭko, Olga V. Golyshina, A. V. Troitskiĭ, K. M. Val'ekho-Roman, R.&mnsp:S. Golovacheva, Tatiana A. Pivovarova: Sulfurococcus yellowstonii sp. nov/--a new species of iron- and sulfur-oxidizing thermoacidophilic Archaeobacterium. ('Sulfurococcus yellowstonii sp. nov. новый вид железо - и сероокисляющей термоацидофильной архебактерии/novyĭ vid zhelezo- i serookisliaiushcheĭ termoatsidofil'noĭ arkhebakterii). In: Микробиология (Mikrobiologiia), Band 63, Nr. 4, Juli-August 1994, S. 668-682, PMID 7845250, Bangor University: Abstract (Abstract:englisch, Studie:russisch).
↑ ab
Adrian Hetzer, Hugh W. Morgan, Ian R. McDonald, Christopher J. Daughney: Microbial life in Champagne Pool, a geothermal spring in Waiotapu, New Zealand. In: Extremophiles. 11. Jahrgang, Nr.4, Juli 2007, S.605–614, doi:10.1007/s00792-007-0073-2, PMID 17426919 (englisch). Siehe insbes. Fig. 4A.
Taxon History. In: GTDB. Abgerufen am 10. Mai 2023 (englisch).
↑
Xi Feng, Yanan Li, Chang Tian, Wei Yang, Xinyu Liu, Changyi Zhang, Zhirui Zeng: Isolation of archaeal viruses with lipid membrane from Tengchong acidic hot springs. In: Frontierrs in Microbiology. Band 14, Sec. Biology of Archaea, Research Topic: Diversity, Ecology and Evolution of Archaeal Viruses, 29. März 2023; doi:10.3389/fmicb.2023.1134935 (englisch)
↑
Monika Häring, Reinhard Rachel, Xu Peng, Roger A. Garrett, David Prangishvili: Viral Diversity in Hot Springs of Pozzuoli, Italy, and Characterization of a Unique Archaeal Virus, Acidianus Bottle-Shaped Virus, from a New Family, the Ampullaviridae. In: Journal of Virology, Band 79, Nr. 15, 1. August 2005, ISSN0022-538X; S. 9904-9911; doi:10.1128/jvi.79.15.9904-9911.2005 (englisch).
↑ abcde
Ruben M. Ceballos, Caleb D. Marceau, Joshua O. Marceau, Steven Morris, Adam J. Clore, Kenneth M. Stedman: Differential virus host-ranges of the Fuselloviridae of hyperthermophilic Archaea: implications for evolution in extreme environments. In: Frontiers in Microbiology, Sec. Extreme Microbiology, Band 3, 24. August 2012; doi:10.3389/fmicb.2012.00295 (englisch).
↑
Blake Wiedenheft, Kenneth Stedman, Francisco Roberto, Deborah Willits, Anne-Kathrin Gleske, Luisa Zoeller, Jamie Snyder, Trevor Douglas, Mark Young: Comparative Genomic Analysis of Hyperthermophilic Archaeal Fuselloviridae Viruses. In: Journal of Virology, Band 58, Nr. 4, 15 Februar 2004; doi:10.1128/jvi.78.4.1954-1961.2004, PMC 369504 (freier Volltext), PMID 14747560 (englisch)
↑
David Prangishvili, Tomohiro Mochizuki, Mart Krupovic, ICTV Report Consortium: ICTV virus taxonomy profile: Guttaviridae. In: Journal of General Virology, Band 99, Nr. 3, Februar 2018; doi:10.1099/jgv.0.001027, ResearchGate:323094322 (englisch). Siehe insbes. Fig. 1 und Fig. 2.
↑
Hans Peter Arnold, Ulrike Ziese, Wolfram Zillig: SNDV, a Novel Virus of the Extremely Thermophilic and Acidophilic Archaeon Sulfolobus. In: Virology, Band 272, Nr. 2, 5. Juli 2000, S. 409-416; doi:10.1006/viro.2000.0375, PMID 10873785 (englisch).
↑
Ariane Bize, Xu Peng, Maria Prokofeva, Kirsty Maclellan, Soizick Lucas, Patrick Forterre, Roger A. Garrett, Elizaveta A. Bonch-Osmolovskaya, David Prangishvili: Viruses in acidic geothermal environments of the Kamchatka Peninsula. In: Research in Microbiology, Band 159, Nr. 5, Juni 2008, S. 358-366; doi:10.1016/j.resmic.2008.04.009, PMID 18565736, Epub 8. Mai 2008 (englisch).
↑
Gisle Vestergaard, Monika Häring, Xu Peng, Reinhard Rachel, Roger A. Garrett, David Prangishvili: A novel rudivirus, ARV1, of the hyperthermophilic archaeal genus Acidianus. In: Virology, Band 336, Ne. 1, 25. Mai 2005, S. 83-92; doi:10.1016/j.virol.2005.02.025, PMID 15866073 (englisch)






.jpg)

{kind=link}