Mikrobialith



Mikrobialithen[4] (englisch microbialites, von neulateinisch microbia ‚Mikrob(i)en‘, ursprünglich altgriechisch μικρός mikros, deutsch ‚klein‘ und βίος bios, deutsch ‚Leben‘) sind benthische Sedimentablagerungen aus Karbonatschlamm (mit einem Partikeldurchmesser weniger als 5 μm), die durch die Vermittlung von Mikroben gebildet werden. Der konstituierende Karbonatschlamm ist eine Art „Automikrit“ (d. h. autochthoner Mikrit oder authigener Karbonatschlamm, englisch authigenic carbonate mud). Das bedeutet, dass er in situ ausfällt und nicht hertransportiert und abgelagert wird. Deshalb kann ein Mikrobialith als eine nach der Dunham-Klassifikation als ein Boundstone angesehen werden, bei dem die Riffbildner Mikroben sind und die Ausfällung der Karbonatpartikel durch diese biotisch induziert (in Gegensatz zu einem Seeigel-Gehäuse (englisch test), einer Muschelschale oder einem Skelett). Mikrobialithen können auch als mikrobielle Matten mit Lithifikationsvermögen[A. 3] definiert werden.[5] Bakterien können sowohl im flachen als auch – mit Ausnahme der phototrophen Cyanobakterien – im tiefen Wasser Karbonat ausfällen. Daher können sich grundsätzlich Mikrobialithen unabhängig vom Sonnenlicht bilden.[6]
Mikrobialithen sind die Grundlage vieler lakustrischer (limnische) Ökosysteme. Beispiele sind:
- das Biosystems des Großen Salzsees in Utah mit seinen Millionen von Zugvögeln,[7]
- der alkalische Kratersee von Alchichica[A. 4] (Mexiko), wo sie die Kinderstube des Axolotl (Ambystoma taylori) und einer Vielzahl von Fischen bilden.
- Im Großen Salzsee (Utah) sind speziell an die hypersalinen Bedingungen angepasste Nematoden mit den dortigen Mikrobialithen assoziiert.[8]
Mikrobialithen waren für die Bildung von präkambrischen und phanerozoischen Kalksteinen in vielen verschiedenen marinen und nicht marinen Umgebungen von großer Bedeutung. Die große Zeit der Stromatolithen (meist von Cyanobakterien gebildete geschichtete Mikrobialithen) war vor 2,8 bis 1,0 Milliarden Jahren, als sie die Hauptbestandteile von Karbonatplattformen waren.[6]
Klassifikation
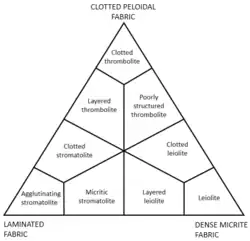
Die Mikrobialithen können nach drei verschiedenen Aufbautypen unterteilt werden:[6]
- stromatolithisch: geschichtet, wobei die Schichten laminiert (miteinander verklebt) oder agglutiniert (verklumpt) sind und so einen Stromatolithen bildet.
- thrombolithisch: mit verklumptem peloidalem Gewebe, wie im petrographischen Mikroskop zu erkennen. Die Dichte der Peloide ist variabel. Das Gestein zeigt eine dendritische Struktur und wird dann als Thrombolith[9][10] bezeichnet.
- leiolitisch: ohne Schichtung und ohne geklumptes Peloidgewebe, nur aus einem dichten Automikrit bestehend und dann als Leiolith[11] bezeichnet.
Evolution
Mikrobialithen spielten eine wichtige Rolle bei den Veränderungen, die die frühe Erdatmosphäre erfassen hat, da sie die ökologische Nischen darstellten, in denen sich die ersten mikrobiellen die Sauerstoff freisetzenden Stoffwechselprozesse herausbilden konnten. Die Mikrobialithen sättigten die marinen Küstensysteme und später die Uratmosphäre mit Sauerstoff und wandelten diesen von einem reduzierten in einen oxidierten Zustand um (Große Sauerstoffkatastrophe Great Oxigenstion Event, auch Great Oxidation Event).[12] Fossile Mikrobialithen aus dem Präkambrium und Phanerozoikum, speziell vom Typ der Stromatolithen, gehören zu den ersten Belegen für Lebensgemeinschaften verschiedener Organismen. Die ältesten Mikrobialithen werden auf mindestens 3,5 Milliarden Jahre datiert.[13] Fossile Belege deuten darauf hin, dass Mikrobialithen produzierende Organismen vom frühen Archaikum bis zum späten Proterozoikum eine sehr häufige Lebensform waren, bis ihre Gemeinschaften durch räuberische Foraminiferen und andere eukaryotische Mikroorganismen zurückgingen.[14] Mikrobialithen traten aber kurz nach dem Massenaussterben an der Perm-Trias-Grenze wieder gehäuft auf.[15][16][17]
Bildung von Mikrobialithen
Die Bildung von Mikrobialithen ist komplex und stellt einen kontinuierlichen Prozess der Ausfällung und (Wieder-)Auflösung dar. Dabei sind verschiedene mikrobielle Stoffwechselvorgänge gekoppelt und es liegt ein hoher Sättigungsindex (SI)[18][19] von Ionen im Wasser vorliegt.[20]
Für Mikrobialithen sind zwei mögliche Entstehungsmechanismen bekannt:
- Mineralische Ausfällung (mineral precipitation): Sie ist der Hauptbildungsprozess von Mikrobialithen und kann durch anorganische Ausfällung oder durch den passiven Einfluss mikrobieller Stoffwechselvorgänge erfolgen. Es kann auch zu einer Ausfällung aufgrund der Sättigung der Mikroumgebung kommen, wenn extrazelluläre polymere Substanzen (EPS) schnell abgebaut werden, was die Ionensättigung erhöht.[21]
- Einfangen und Binden (trapping and binding): Wenn die mikrobielle Gemeinschaft mineralische Partikel aus der Umgebung enthält, die an den extrazellulären polymeren Substanzen (EPS) haften. Dieser Prozess ist heute ziemlich vorherrschend, er wurde beispielsweise in den modernen Mikrobialithen der Shark Bay (Australien) und der Bahamas beschrieben. Es hat sich aber gezeigt, dass er in der 3,5 Milliarden Jahre langen geologischen Geschichte der Mikrobialithen sehr selten vorkam.[22]
Vorkommen moderner Mikrobialithen
Lebende oder allgemein „moderne“ Mikrobialithen (weniger als 20.000 Jahre alt) sind selten und finden sich nur an einigen Orten wie:
- Kraterseen:
- Blue Lake alias Wa(a)rwar (Südaustralien),[23] Satonda-See (Indonesien), Dziani Dzaha (Mayotte), Maarsee von Alchichica[A. 5] (Mexiko), Vai Lahi[24][25] mit Vai Sii alias Vai Mafaʻaho[26] (beide Niuafoʻou, Tonga), Salda-See (Türkei)
- Salinen, hypersaline Seen und Lagunen:
- Pyramid Lake und Großer Salzsee (USA), Vansee (Türkei), Laguna Brava[27][28] und Lagune Tebenquiche alias Tebinquiche[29][30][31] (Chile)
- Natronseen (Sodaseen):
- Lake Thetis[32][33][34] (Westaustralien), Sarmiento-See (Chile),[35][36] Nuo'ertu-See[37][38][39][40] und Huhejaran-See alias Huhe-Jilin-See[39][37] (beide China), Mono Lake (USA), Turkana-See (Kenia), Petuchowskoje-See alias Petuchowo-See[41][42][43] (Altai).
- Süßwasserseen/Lagunen:
- Lagoa Salgada[44] (Brasilien), Laguna Negra[45] (Catamarca, Argentinien), Lagunas de Ruidera[46] (Spanien), Lagune von Bacalar (Mexiko), Lake Richmond (Australien), Pavilion Lake (Kanada), Green Lake (USA)
- Meere/Flussmündungen/Ästuare:
- Shark Bay (Westaustralien), Highbourne Cay[A. 6] (Bahamas), Tikehau (Französisch-Polynesien), Cayo Coco (Kuba), Lake Clifton (Westaustralien).[48]
Die lebenden Mikrobialithen werden auch manchmal etwas ungenau als „lebende Steine“ bezeichnet,[10] diese Begrifflichkeit ist aber auch für verschiedene Vertreter der Mittagsblumengewächse (Aizoaceae) gängig, wie z. B. Lapidaria margaretae; wird aber auch gelegentlich für Moqui-Marbles verwendet (siehe Begriffsklärungsseite Lebende Steine).
Zusammensetzung
Mikrobialithen bestehen aus Schichten einer organischen Komponente und eines Minerals.[49] Die organische Komponente ist eine speziell organisierte mikrobielle Matte, in der Gemeinschaften (Konsortien) von Mikroorganismen mit unterschiedlichen Metabolismen interagieren und Mikro-Nische bilden, in der aerobe und anaerobe, oft (aber nicht notwendig) phototrophe Organismen nebeneinander existieren: Stickstofffixierer, Schwefel-Reduzierer, Methanotrophe, Methanogene, Eisen-Oxidierer und eine Unzahl heterotropher Zersetzer.[50] Die mineralische Komponente besteht aus Karbonaten, in der Regel Kalziumkarbonat- oder Magnesiumkarbonat-Mineralien wie Hydromagnesit, aber auch durch Sintern (Verdichten) entstandene Silikate. Sie enthält mineralische Formen von Schwefel, Eisen (Pyrit, FeS2) oder Phosphor.[21] Karbonat ist in der Regel eine Art autogener Automikrit, d. h. es wird in situ ausgefällt. Mikrobialithen können als eine Art biogenes Sedimentgestein betrachtet werden, bei dem die Riffbildner Mikroben sind und die Karbonatausfällung durch diese herbeigeführt (induziert) wird. Die Mikroorganismen können Karbonat sowohl in flachen als auch in tiefen Gewässern ausfallen lassen.
Mikrobialith-assoziierte Mikroorganismen
Die Vielfalt der alleine an der Oberfläche von Mikrobialithen lebenden Organismen wurde in zahlreichen Studien untersucht.[51][52] Diese Vielfalt ist häufig sehr groß, sie umfasst Bakterien, Archaeen und Mikroeukaryoten. Während die phylogenetische Vielfalt dieser mikrobiellen Gemeinschaften mit Hilfe der Molekularbiologie im Allgemeinen recht gut untersucht ist, bleibt die Identität der Organismen, die zur Karbonatbildung beitragen, unklar (Stand 2021). Interessanterweise gibt es einige Mikroorganismen, die in den Mikrobialithen verschiedener Seen vorkommen, sie definieren daher ein „Kernmikrobiom“ (core microbiome).[53][51] Bei den Karbonat ausfällenden und so die Mikrobialithen aufbauenden Mikroben handelt es sich meist um Prokaryoten (Bakterien und Archaeen). Die bekanntesten karbonatbildenden Bakterien sind Cyanobakterien und sulfatreduzierende Bakterien (sulfate reducing bacteria, SRBs).[20] Es können aber auch andere Bakterien eine wichtige Rolle spielen, wie z. B. Bakterien, die anoxygene Photosynthese betreiben.[54] Archaeen sind häufig extremophil, d. h. leben in entlegenen Umgebungen, in denen andere Organismen nicht leben können, wie z. B. die weißen Raucher am Grund der Ozeane. Eukaryotische Mikroben (Mikroeukaryoten) produzieren weniger Karbonat als diese Prokaryoten und leisten daher nur einen geringen Beitrag zu Aufbau der Mikrobialithen.[55]
Relevanz
In der Paläontologie besteht großes Interesse an der Untersuchung fossiler Mikrobialithen, da sie als bioklimatische Indikatoren fungieren und wichtige Daten über das Paläoklima liefern.[56] Da sie zu den ersten Lebensformen auf der Erde gehören, besteht auch in der Astrobiologie Interesse an ihrer Untersuchung, für den Fall, dass man auf anderen Planeten[A. 7] in unserem Sonnensystem diese Strukturen finden sollte.[57] Die Untersuchung von modernen Mikrobialithen kann relevante Informationen liefern, indem sie als Umweltindikatoren für die Bewirtschaftung und Erhaltung von Naturschutzgebieten dienen.[58] Aufgrund ihrer Fähigkeit, Mineralien zu bilden und detritisches Material auszufällen, wurden biotechnologische und bioremediale Anwendungen in aquatischen Systemen zur Kohlendioxidbindung vorgeschlagen, so dass Mikrobialithen als Kohlenstoffsenken (genauer: Kohlendioxidsenke) fungieren können.[59]
Anmerkungen
- ↑ neu gezeichnet und vereinfacht nach Schmid (1996)
- ↑ Strelley-Pool-Hornstein ist englisch Strelley Pool Chert (SPC)
- ↑ der Eigenschaft, per Fossilisation bzw. Diagenese eine Versteinerung bilden zu können
- ↑ 19,4167° N, 97,4° W, in Puebla (Mexiko)
- ↑ Laguna de Alchichica, englisch Lake Alchichica 19,4167° N, 97,4° W, Zentralmexiko
- ↑ Das Highbourne Cay (Cay ist die örtliche Bezeichnung für Koralleninseln, vgl. Key wie Florida Keys und Cayo Coco) ist eine Koralleninsel im Norden der Exuma Cays.
- ↑ oder den Monden der Gasriesen
Weblinks und Literatur
- Bacterial Layers: Living Fossils: Microbialites. Living Fossils: Microbialites. Digital Atlas of Ancient Life (digitalatlasofancientlife.org) der Paleontological Research Institution (englisch).
Einzelnachweise
- ↑ Dieter U. Schmid: Mikrobolithe und Mikroinkrustierer aus dem Oberjura. In: Profil. 9. Jahrgang, 1996, S. 101–251.
- ↑ Strelley Pool. Auf Mapcarta (de).
- ↑ Martin Julian Van Kranendonk: Pilbara Craton. In: Encyclopedia of Astrobiology (Eintrag im Nachschlagwerk), Springer Nature Link, S. 1257–1260, Print-ISBN 978-3-642-11271-3; doi:10.1007/978-3-642-11274-4_1213, Online-ISBN 978-3-642-11274-4 (englisch).
- ↑ Immo Schülke, Stefan Delecat, Carsten Helm: Oberjura-Riffe in NW-Deutschland: Ein Überblick (PDF). In: Mitteilungen des geologischen Instituts der Universität Hannover (Institut für Geologie und Paläontologie der Universität Hannover), Band 38, ISSN 0440-2812, Juli 1998, S. 191–202, 1 Abb., 1 Tab.
- ↑ Christophe Dupraz, Pieter T. Visscher: Microbial lithification in marine stromatolites and hypersaline mats. In: Trends in Microbiology. 13. Jahrgang, Nr. 9, September 2005, S. 429–438, doi:10.1016/j.tim.2005.07.008, PMID 16087339 (englisch).
- ↑ a b c Erik Flügel, mit einem Beitrag von Axel Munnecke: Microfacies of Carbonate Rocks: Analysis, Interpretation and Application. 2. Auflage. Springer, Heidelberg 2010, ISBN 978-3-642-03795-5, ISBN 978-3-642-03796-2, 984 bzw. 1007 Seiten (englisch).
- ↑ Hollie Brown, Mark Milligan et al.: Drought Negatively Impacting Great Salt Lake Microbialites and Ecosystem. Auf: Utah Geological Survey. Department of Natural Resources (DNR), 15. Juli 2021 (englisch). Memento im Webarchiv vom 24. März 2025.
- ↑
Julie Jung, Tobias Loschko, Shelley Reich, Maxim Rassoul-Agha, Michael S. Werner: Newly identified nematodes from the Great Salt Lake are associated with microbialites and specially adapted to hypersaline conditions. In: Proceedings of the Royal Socirety B, Band 291, Nr. 2018, 13. März 2024; Online-ISSN 1471-2954, doi:10.1098/rspb.2023.2653, PMID 38471558 (englisch). Dazu:
- With discovery of roundworms, Great Salt Lake’s imperiled ecosystem gets more interesting. Auf EurekAlert! vom 13. März 2024. Quelle: University of Utah.
- Carly Cassella Scientists Thought Only Two Animals Lived in The Great Salt Lake. They Were Wrong. Auf sciencealert vom 17. März 2024.
- ↑ Wikidata: Thrombolithen (Q7798340).
- ↑ a b
Lake Clifton Thrombolites. Auf: westernaustralia.com (englisch). Dazu:
- Petra Paul: Thrombolithen – Lebende Steine von Lake Clifton: Sprungbrett in die Evolution!. Auf Passenger on Earth (passengeronearth.com) vom 3. Februar 2012.
- ↑ Leiolite. A rock subtype. MinDat, Hudson Institute of Mineralogy. Stand: 9. Januar 2025 (englisch).
- ↑ Bernard Laval, Sherry L. Cady, John C. Pollack, Christopher P. McKay, John S. Bird, John P. Grotzinger, Derek C. Ford, Harry R. Bohm: Modern freshwater microbialite analogues for ancient dendritic reef structures. In: Nature. 407. Jahrgang, Nr. 6804, Oktober 2000, ISSN 0028-0836, S. 626–629, doi:10.1038/35036579, PMID 11034210, bibcode:2000Natur.407..626L (englisch).
- ↑ Stanley M. Awramik: Precambrian Columnar Stromatolite Diversity: Reflection of Metazoan Appearance. In: Science. 174. Jahrgang, Nr. 4011, 19. November 1971, ISSN 0036-8075, S. 825–827, doi:10.1126/science.174.4011.825, PMID 17759393, bibcode:1971Sci...174..825A (englisch).
- ↑ Joan M. Bernhard, Virginia P. Edgcomb, Pieter T. Visscher, Anna McIntyre-Wressnig, Roger E. Summons, Mary L. Bouxsein, Leeann Louis, Marleen Jeglinski: Insights into foraminiferal influences on microfabrics of microbialites at Highborne Cay, Bahamas. In: Proceedings of the National Academy of Sciences. 110. Jahrgang, Nr. 24, 28. Mai 2013, ISSN 0027-8424, S. 9830–9834, doi:10.1073/pnas.1221721110, PMID 23716649, PMC 3683713 (freier Volltext), bibcode:2013PNAS..110.9830B (englisch).
- ↑ Arnaud Brayard, Emmanuelle Vennin, Nicolas Olivier, Kevin G. Bylund, Jim Jenks, Daniel A. Stephen, Hugo Bucher, Richard Hofmann, Nicolas Goudemand, Gilles Escarguel: Transient metazoan reefs in the aftermath of the end-Permian mass extinction. In: Nature Geoscience. 4. Jahrgang, Nr. 1, 18. September 2011, S. 693–697, doi:10.1038/ngeo1264, bibcode:2011NatGe...4..693B (englisch).
- ↑ Siqi Wu, Zhong-Qiang Chen, Yuheng Fang, Yu Pei, Hao Yang, James Ogg: A Permian-Triassic boundary microbialite deposit from the eastern Yangtze Platform (Jiangxi Province, South China): Geobiologic features, ecosystem composition and redox conditions. In: Palaeogeography, Palaeoclimatology, Palaeoecology. 486. Jahrgang, 15. November 2017, S. 58–73, doi:10.1016/j.palaeo.2017.05.015, bibcode:2017PPP...486...58W (englisch).
- ↑ Steve Kershaw, Sylvie Crasquin, Yue Li, Pierre-Yves Collin, Marie-Béatrice Forel, X. Mu, Aymon Baud, Yongbiao Wang, Shucheng Xie, Florian Maurer, Li Guo: Microbialites and global environmental change across the Permian–Triassic boundary: a synthesis. In: Geobiology. 10. Jahrgang, Nr. 1, 13. November 2011, ISSN 1472-4677, ResearchGate:51793045, S. 25–47, doi:10.1111/j.1472-4669.2011.00302.x (englisch).
- ↑ Wiktionary: Sättigung.
- ↑ Sättigungsindex. Lexikon der Geowissenschaften (spektrum.de).
- ↑ a b Anderson A. P. Chagas, Gregory E. Webb, Robert V. Burne, Gordon Southam: Modern lacustrine microbialites: Towards a synthesis of aqueous and carbonate geochemistry and mineralogy. In: Earth-Science Reviews. 162. Jahrgang, November 2016, ISSN 0012-8252, S. 338–363, doi:10.1016/j.earscirev.2016.09.012, bibcode:2016ESRv..162..338C (englisch).
- ↑ a b Christophe Dupraz, R. Pamela Reid, Olivier Braissant, Alan W. Decho, R. Sean Norman, Pieter T. Visscher: Processes of carbonate precipitation in modern microbial mats. In: Earth-Science Reviews. 96. Jahrgang, Nr. 3, Oktober 2009, ISSN 0012-8252, S. 141–162, doi:10.1016/j.earscirev.2008.10.005, bibcode:2009ESRv...96..141D (englisch).
- ↑ Pablo Suarez-Gonzalez, M. Isabel Benito, I. E. Quijada, Ramón Mas, Sonia Campos-Soto: ‘Trapping and binding’: A review of the factors controlling the development of fossil agglutinated microbialites and their distribution in space and time. In: Earth-Science Reviews. 194. Jahrgang, 1. Juli 2019, ISSN 0012-8252, hdl:10651/56917, ResearchGate:332997220, S. 182–215, doi:10.1016/j.earscirev.2019.05.007 (englisch).
- ↑ Wikidata: Blue Lake / Warwar (Q2790756).
- ↑ Wikidata: Vai Lahi (Q16902456).
- ↑ Vai Lahi (crater lake). Auf GeoNames.
- ↑ Vaii Sii (crater lake). Auf GeoNames.
- ↑ Maria Cecilia Rasuk, Pieter T. Visscher, Manuel Contreras, María Eugenia Farías: Mats and Microbialites from Laguna La Brava. In: María Eugenia Farías (Hrsg.): Microbial Ecosystems in Central Andes Extreme Environments, Mai 2020, S. 221–230; doi:10.1007/978-3-030-36192-1_15, ResearchGate:341692112 (englisch).
- ↑ Laguna Brava, Chile » Antofagasta. Auf GeoNames. Anm.: Südlich des Salar de Atacama, nicht zu verwechseln mit Sierra Nevada de Lagunas Bravas.
- ↑ Ana B. Fernandez, Maria C. Rasuk, Pieter T. Visscher, Manuel Contreras, Fernando Novoa, Daniel G. Poire, Molly M. Patterson, Antonio Ventosa, Maria E. Farias: Microbial Diversity in Sediment Ecosystems (Evaporites Domes, Microbial Mats, and Crusts) of Hypersaline Laguna Tebenquiche, Salar de Atacama, Chile. In: Frontiers in Microbiology, Sec. Extreme Microbiology, Band 7, 22. August 2016; doi:10.3389/fmicb.2016.01284 (englisch).
- ↑ Laguna Tebinquiche. Auf GeoNames.
- ↑ Martin Mergili: Laguna Tebinquinche (sic!). Stand: 26. Juli 2020 (anden.at). Anm.: Der Name Tebenquiche ist möglicherweise verschrieben zu Tebinquinche.
- ↑ Wikidata: Lake Thetis (Q6478020).
- ↑ Lake Thetis, nahe (östlich) von Cervantes. Auf Mapcarta (de).
- ↑ Lake Thetis, Australia. Auf GeoNames.
- ↑ Wikidata: Sarmiento-See (Q2477323).
- ↑ Lago Sarmiento de Gamboa, Chile. Auf GeoNames.
- ↑ a b Badain Jaran. Der See 诺尔图 Nuo'ertu ist der zweite südöstlich von der Marke für die Badain-Jaran-Wüste, der Huhejaran-See befindet sich direkt im Norden, hier als Huhe-Jilin bezeichnet. Auf GeoNames.
- ↑ 巴丹吉林嘎查 (Dorf Badan Jilin Gacha). Der Nuo'ertu-See befindet sich südöstlich vom angezeigten Dorf. Auf Mapcarta (de).
- ↑ a b Huhejaran-See (Gewässer). Auf GeoFinder (geofinder.ch). Der Nuo'ertu-See ist der hier nicht benannte See etwas direkt im Süden, südwestlich von 巴丹吉林嘎查 ‚Badan Jilin Gacha‘.
- ↑ Ricardo Silva-Rweisswald: full view nuoertu lake. Auf EyeEm (eyeem.com).
- ↑ Olga S. Samylina, Lyubov V. Zaytseva: Characterization of modern dolomite stromatolites from hypersaline Petukhovskoe Soda Lake, Russia. In: Scandinavian University Press, Band 52, Nr. 1, 1. August 2018, S. 1–13; doi:10.1111/let.12286 (englisch).
- ↑ ozero Petukhovskoe, Region Altai, Russland. W-SW von Severskiy. Auf Bing Maps.
- ↑ Ozero Petukhovo (Озеро Петухово). W-SW von Severka. Auf GeoNames.
- ↑ Wikidata: Lagoa Salgada (Q973069).
- ↑ Wikidata: Laguna Negra (Q99539201).
- ↑ Wikidata: Lagunas de Ruidera (Q2271809).
- ↑ Wikidata: Clinton Creek, Siedlung in Kanada (Q2979349).
- ↑ Wikidata: Lake Clifton (Q6475415).
- ↑ Carla M. Centeno, Pierre Legendre, Yislem Beltrán, Rocío J. Alcántara-Hernández, Ulrika E. Lidström, Matthew N. Ashby, Luisa I. Falcón: Microbialite genetic diversity and composition relate to environmental variables. In: FEMS Microbiology Ecology. 82. Jahrgang, Nr. 3, 2. August 2012, ISSN 0168-6496, S. 724–735, doi:10.1111/j.1574-6941.2012.01447.x, PMID 22775797 (englisch).
- ↑ Richard Allen White III, Amy M. Chan, Gregory S. Gavelis, Brian S. Leander, Allyson L. Brady, Gregory F. Slater, Darlene S. S. Lim, Curtis A. Suttle: Metagenomic Analysis Suggests Modern Freshwater Microbialites Harbor a Distinct Core Microbial Community. In: Frontiers in Microbiology. 6. Jahrgang, 28. Januar 2016, ISSN 1664-302X, S. 1531, doi:10.3389/fmicb.2015.01531, PMID 26903951, PMC 4729913 (freier Volltext), bibcode:2016FrMic...3.1531W (englisch).
- ↑ a b Miguel Iniesto, David Moreira, Guillaume Reboul, Philippe Deschamps, Karim Benzerara, Paola Bertolino, Aurélien Saghaï, Rosaluz Tavera, Purificación López‐García: Core microbial communities of lacustrine microbialites sampled along an alkalinity gradient. In: Environmental Microbiology. 23. Jahrgang, Nr. 1, Januar 2021, ISSN 1462-2912, S. 51–68, doi:10.1111/1462-2920.15252, PMID 32985763 (englisch).
- ↑ Estelle Couradeau, Karim Benzerara, David Moreira, Emmanuelle Gérard, Józef Kaźmierczak, Rosaluz Tavera, Purificación López-García: Prokaryotic and Eukaryotic Community Structure in Field and Cultured Microbialites from the Alkaline Lake Alchichica (Mexico). In: PLOS ONE. 6. Jahrgang, Nr. 12, 14. Dezember 2011, ISSN 1932-6203, S. e28767, doi:10.1371/journal.pone.0028767, PMID 22194908, PMC 3237500 (freier Volltext), bibcode:2011PLoSO...628767C (englisch).
- ↑ Richard Allen White, Ian M. Power, Gregory M. Dipple, Gordon Southam, Curtis A. Suttle: Metagenomic analysis reveals that modern microbialites and polar microbial mats have similar taxonomic and functional potential. In: Frontiers in Microbiology. 6c. Jahrgang, 23. September 2015, ISSN 1664-302X, S. 966v, doi:10.3389/fmicb.2015.00966, PMID 26441900, PMC 4585152 (freier Volltext) – (englisch).
- ↑ Aurélien Saghaï, Yvan Zivanovic, Nina Zeyen, David Moreira, Karim Benzerara, Philippe Deschamps, Paola Bertolino, Marie Ragon, Rosaluz Tavera, Ana I. López-Archilla, Purificación López-García: Metagenome-based diversity analyses suggest a significant contribution of non-cyanobacterial lineages to carbonate precipitation in modern microbialites. In: Frontiers in Microbiology. 6. Jahrgang, 5. August 2015, ISSN 1664-302X, S. 797, doi:10.3389/fmicb.2015.00797, PMID 26300865, PMC 4525015 (freier Volltext) – (englisch).
- ↑ Robert Riding: Microbial carbonates: the geological record of calcified bacterial–algal mats and biofilms. In: Sedimentology. 47. Jahrgang, s1, 5. Januar 2000, ISSN 1365-3091, S. 179–214, doi:10.1046/j.1365-3091.2000.00003.x (englisch).
- ↑ Gregory E. Webb, Balz S. Kamber: Rare earth elements in Holocene reefal microbialites: a new shallow seawater proxy. In: Geochimica et Cosmochimica Acta. 64. Jahrgang, Nr. 9, Mai 2000, ISSN 0016-7037, S. 1557–1565, doi:10.1016/s0016-7037(99)00400-7, bibcode:2000GeCoA..64.1557W (englisch).
- ↑ Nora Noffke: Ancient Sedimentary Structures in the <3.7 Ga Gillespie Lake Member, Mars, That Resemble Macroscopic Morphology, Spatial Associations, and Temporal Succession in Terrestrial Microbialites. In: Astrobiology. 15. Jahrgang, Nr. 2, Februar 2015, ISSN 1531-1074, S. 169–192, doi:10.1089/ast.2014.1218, PMID 25495393, bibcode:2015AsBio..15..169N (englisch).
- ↑ Joachim Reitner, Volker Thiel (Hrsg.): Encyclopedia of geobiology. Springer, Dordrecht 2011, ISBN 978-1-4020-9212-1, doi:10.1007/978-1-4020-9212-1 (englisch).
- ↑ Ranieri Cancedda, Andrea Banfi, Gianni Ciofani, Dan (Annie) Du (Hrsg.): Frontiers in Bioengineering and Biotechnology. Frontiers Media SA, doi:10.3389/fbioe (englisch).