Lambdotherium
| Lambdotherium | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|

Teilskelett von Lambdotherium | ||||||||||||
| Zeitliches Auftreten | ||||||||||||
| Unteres Eozän (Wasatchium, Lostcabinium) | ||||||||||||
| 53 bis 52,1 Mio. Jahre | ||||||||||||
| Fundorte | ||||||||||||
| ||||||||||||
| Systematik | ||||||||||||
| ||||||||||||
| Wissenschaftlicher Name | ||||||||||||
| Lambdotherium | ||||||||||||
| Cope, 1880 | ||||||||||||
Lambdotherium ist eine Gattung aus der ausgestorbenen Familie der Lambdotheriidae, welche als nahe mit den Brontotheriidae verwandt gilt und manchmal auch innerhalb dieser eingegliedert wird. Funde beschränken sich weitgehend auf Gebissteile und einige wenige partiell erhaltene Skelette. Sie wurden im westlichen Nordamerika in den US-Bundesstaaten Wyoming und Colorado gefunden. Mit einer Datierung in das Untere Eozän vor gut 53 bis 52 Millionen Jahren gehören sie zu den ältesten Nachweisen aus der Verwandtschaftsgruppe der Brontotherien. Es handelt sich um vergleichsweise kleine Tiere mit einem für Unpaarhufer ursprünglichen Schädelbau. Allerdings war das Gebiss bereits leicht reduziert. Zudem zeigt sich an den oberen Mahlzähnen die für die Brontotherien typische Gestaltung mit einer W-förmig verlaufenden Schmelzleiste. Die Gattung wurde im Jahr 1880 wissenschaftlich eingeführt. In der Regel ist eine Art anerkannt. Der Verweis in die Verwandtschaftsgruppe der Brontotherien war forschungsgeschichtlich nicht immer gegeben, da einige Autoren eine Beziehung zu den Palaeotheriidae und damit zum Umfeld der Pferde vermuteten.
Merkmale
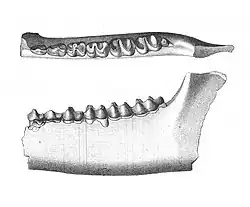
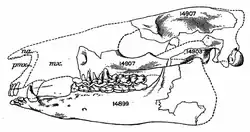
Der überwiegende Teil des bekannten Fundmaterials von Lambdotherium besteht aus Gebissresten, vollständige Schädel oder Teile des postcranialen Skeletts sind eher selten. Ein einzelner, vollständig überlieferter Schädel maß 18,2 cm in der Länge und 9,8 cm in der Breite ermittelt an den Jochbögen. Im Bereich der Augenfenster wurde er 6,7 cm breit, schnürte aber hinter diesen auf rund 3,3 cm Breite ein. Der Gesichtsschädel und der Hirnschädel waren mit Längen von 8,5 beziehungsweise 8,0 cm nahezu gleich lang. Dies ist untypisch für die meisten Brontotherien, bei denen der hintere Schädelabschnitt den vorderen deutlich an Länge übertraf. Dadurch zeigte sich bei Lambdotherium die Rostrum im Vergleich zu den späteren Brontotherien noch relativ langgestreckt, zudem wies sie eine röhrenartige Gestaltung auf. In seiner generellen Form erinnerte der Schädel an jenen eines ursprünglichen Unpaarhufers. Dies spiegelt sich im Verlauf der Stirnlinie in Seitenansicht wider, die leicht konvex aufgewölbt war, während sie bei den stammesgeschichtlich jüngeren Brontotherien charakteristisch eindellte. Auch der kleine Naseninnenraum, der auf Höhe des Eckzahns endete, entspricht den Verhältnissen bei den frühen Unpaarhufern. Das Foramen infraorbitale lag oberhalb des dritten Prämolaren, öffnete sich nach vorn und war klein. Die Orbita befand sich auf Höhe des ersten und zweiten Molaren und besaß einen Durchmesser von 2,6 cm. Nur wenig weiter vorgeschoben setzte der vordere Abschnitt des Jochbogens an. Der Hirnschädel wurde von einem kräftigen Scheitelkamm überragt. Er reichte rückwärts bis zum Hinterhauptsbein, das in Aufsicht schmal war und in Seitenansicht nach hinten überhing. Der Unterkiefer war allgemein schlank gebaut, seine Höhe unterhalb des zweiten Molaren betrug gut 3,2 cm. Ein markantes Kennzeichen bildete die lang ausgezogene Symphyse am vorderen Ende. Der Kronenfortsatz ragte steil auf.[1][2][3]
Das Gebiss bestand aus 42 Zähnen mit folgender Zahnformel: Im Unterkiefer fehlte somit der vorderste Prämolar, wodurch Lambdotherium von anderen frühen Brontotherien wie Palaeosyops abweicht, die noch über die vollständige Bezahnung der Höheren Säugetiere verfügten. Dies bedingte auch ein ausgeprägteres Diastema zwischen dem vorderen und hinteren Gebiss in der unteren Zahnreihe als bei anderen frühen Brontotherien, es bemaß sich auf gut 2,4 cm. In der oberen Zahnreihe bestanden einige kürzere Zahnlücken von gut 0,5 bis 1,2 cm Ausdehnung hinter dem letzten Schneidezahn und dem ersten Prämolaren, was zu einer isolierten Position des Eckzahns führte. Die oberen und unteren Schneidezähne waren meißel- bis spatelartig geformt und bildeten eine halbkreisförmige Anordnung. Die Eckzähne der jeweiligen oberen und unteren Gebisshälfte wiesen eine deutliche Spitze auf, wobei der untere markanter seitlich verschmälert und nach hinten gebogen war. Die Prämolaren zeigten sich mit Ausnahme des letzten unteren nur wenig molarisiert, glichen also kaum den Molaren, Sie charakterisierte ein einzelner Haupthöcker, der Protoconus (oben) oder das Protoconid (unten), welcher jeweils von mehr oder weniger ausgeprägten Nebenhöckern begleitet wurde. Die Mahlzähne besaßen im Oberkiefer eine auffallend W-förmige Kauoberflächengestaltung als typisches Kennzeichen der Brontotherien. Hervorgerufen wurde diese durch die Scherleiste des Ectolophs, die zwischen den wangenseitigen Haupthöckern (Paraconus und Metaconus) verlief. Auf der Zungenseite erhoben sich die beiden anderen Haupthöcker (Protoconus und Hypoconus) zuzüglich eines kräftigen Nebenhöckers (Paraconul) relativ isoliert. Ähnlich strukturiert waren die unteren Backenzähne, wobei die Haupthöcker hier durch das Protoconid und Metaconid auf dem vorderen (Trigonid) sowie das Hypoconid und Entoconid auf dem hinteren Zahnabschnitt (Talonid) gebildet wurden. Diese verbanden V- oder Λ-förmige Schmelzleisten. Durch den Bau wirkten die unteren Molaren eher lophodont (mit quergestellten Zahnschmelzleisten). Die Länge der unteren Zahnreihe vom zweiten Prämolaren bis zum letzten Molaren variierte zwischen 6,9 und 7,4 cm. Dies ist deutlich weniger als bei anderen frühen Brontotherien wie Eotitanops mit 7,8 bis 10,5 cm oder Palaeosyops mit 14,6 bis 17,8 cm. Die gemittelten Werte für die entsprechende obere Zahnreihe lauten 6,6, 9,1 und 14,4 cm.[2][3][4][5]

Das Körperskelett ist nur ungenügend bekannt. Der Vorderfuß war einzelnen Funden zufolge vierzehig gebaut, was den ursprünglichen Verhältnissen bei den Unpaarhufern entspricht. Allerdings war der äußere Strahl (V) verkürzt und der mittlere (III) offensichtlich noch nicht vergrößert, so dass der Eindruck eines iso-tridactylen Vorderfußes entstand (mit drei gleichlangen Zehen).[3]
Fundstellen

Funde von Lambdotherium sind bisher nur aus Nordamerika bekannt. Der Großteil der Fossilreste stammt aus dem westlichen Bereich der USA, wobei das Hauptfundgebiet im US-Bundesstaat Wyoming liegt. Dort verteilt es sich auf mehrere Gesteinsformationen, von denen die Wind-River-, die Wasatch-, die Willwood- und die Green-River-Formation von größerer Bedeutung sind. Das Typus-Material wurde in der Wind-River-Formation im Wind-River-Becken des west-zentralen Teils von Wyoming entdeckt und umfasst einen Unterkiefer. Von dort stammen aber auch noch weitere Gebissfunde zuzüglich einzelner isolierter Zähne.[6][7][3][8] Nördlich davon im Bighorn-Becken kamen einzelne Zähne und Gebissfragmente aus der Willwood-Formation zu Tage.[9][10] Die Wasatch-Formation lieferte dem gegenüber einige gut erhaltene Schädelteile,[2] darunter auch den bisher vollständigsten Schädel, der im Bridger-Becken im südwestlichen Teil von Wyoming aufgefunden wurde.[11][1] Noch weiter im Südwesten verzahnt die Wasatch-Formation deutlich mit der Green-River-Formation, enthält aber auch hier zumeist einzelne Zähne von Lambdotherium.[12][13] Die Green-River-Formation wiederum ist eine sehr fossilreiche Gesteinsformation und geht auf drei ehemalige Seen zurück. Aus dem westlichsten davon, dem Fossil-Becken, wurden aus dem Fossil Butte Member zwei Teilskelette berichtet. Das eine umfasst den Schädel und den Schultergürtel, das andere den vorderen Rumpfabschnitt. Sie fanden sich mit einer Lage für ersteres im 18-inch layer, für letzteres in den sandwich beds in jeweils unterschiedlicher stratigraphischer Position.[14] Außerhalb dieses zentralen Fundgebietes wurden Reste von Lambdotherium in Form einzelner Unterkieferfragmente aus der Huerfano-Formation im Süden von Colorado berichtet.[15][16]
Alle Funde sind dem Unteren Eozän zuzuweisen. Lambdotherium bildet hierbei die Referenzform des sogenannten Lostcabiniums, einer Stufe in der nordamerikanischen Landsäugetier-Biostratigraphie (NALMA = North American Land Mammal Ages). Das Lostcabinium wiederum nimmt den oberen Abschnitt der lokalen nordamerikanischen Faunenstufe des Wasatchiums ein, welches teilweise synchron zum Unteren Eozän läuft. Die Datierungsansätze für das Lostcabinium liegen im Zeitraum von vor 53 bis 52 Millionen Jahren. Benannt wurde es nach dem gleichnamigen Schichtglied der Wind-River-Formation.[17][18][8]
Paläobiologie
Die Struktur der Backenzähne lässt darauf schließen, dass Lambdotherium sich von gröberer Vegetation wie Blättern und Zweigen ernährte.[14]
Systematik
Innere Systematik der frühen Brontotheriidae nach Mihlbachler 2008[19]
|
Lambdotherium ist eine Gattung aus der Ordnung der Unpaarhufer(Perissodactyla). Innerhalb dieser wird sie teilweise der ausgestorbenen Familie der Lambdotheriidae zugewiesen, die aber mitunter auch als Basalgruppe der ebenfalls erloschenen Familie der Brontotheriidae (ursprünglich Titanotheriidae) gelten und dann als deren früher Abzweig eingestuft werden. Beide Gruppen stehen aufgrund bestimmter zahnmorphologischer Merkmale in einer entfernteren Beziehung zu den heutigen Pferden und gehören damit der übergeordneten Gruppe der Hippomorpha an. Die engere Verwandtschaft der Lambdotheriidae und der Brontotheriidae wird durch die Überfamilie der Brontotherioidea ausgedrückt. Ihr Auftreten ist jeweils auf das Eozän beschränkt, wobei die Lambdotherien nur in dessen unteren Abschnitt vorkamen, die Brontotherien hingegen im gesamten Zeitraum nachweisbar sind. Fossilfunde liegen für beide Gruppen sowohl aus Nordamerika als auch aus Eurasien vor. Die Lambdotherien zeigen in ihrer Anatomie Übereinstimmungen zu den frühen Pferden. Dies ist weitgehend auch für die frühen Brontotherien zutreffend, doch entwickelten diese im Verlauf ihrer Stammesgeschichte teils riesige Vertreter wie etwa Megacerops, deren Charakteristikum knöcherne Hornbildungen auf der Nase sind. Als nahe verwandte Form von Lambdotherium kann Danjiangia aus Ostasien angesehen werden.[20][21][19][22][23]

Die wissenschaftliche Erstbeschreibung von Lambdotherium wurde von Edward Drinker Cope im Jahr 1880 durchgeführt. Er verwendete hierfür ein linkes Unterkieferfragment, an dem noch die Zahnreihe vom zweiten Prämolar bis zum dritten Molar anhaftete (Exemplarnummer AMNH 4863). Es stammt aus Ablagerungen des Unteren Eozäns der Wind-River-Formation im west-zentralen Teil des US-Bundesstaates Wyoming. Der Name der Gattung leitet sich vom griechischen Buchstaben Lambda (λάβδα, Kürzel Λ) und dem Wort θηρίον (thērion) für „Tier“ ab. Er bezieht sich auf die Λ-förmig angeordneten Schmelzleisten auf den unteren Molaren. Gemeinsam mit der Gattung führte Cope die Art L. popoagicum ein, wobei das Artepitheton auf den Popo Agie River, einen Nebenfluss des Wind River verweist.[24][3] Daneben wurden im Laufe der Zeit noch weitere Arten eingeführt, so etwa das von Frederic Brewster Loomis im Jahr 1907 aufgestellte L. primaevum[9] sowie die von Henry Fairfield Osborn im Jahr 1913 benannten Formen L. priscum und L. progressum,[7] beziehungsweise das vom gleichen Autor sechs Jahre später etablierte L. magnum.[15] Ygnacio Bonillas setzte diese jedoch im Jahr 1936 aufgrund weniger zahnmorphologischer Abweichungen mit der Nominatform gleich und erklärte die bestehenden Größenvariationen zwischen den einzelnen Formen als intraspezifisch.[2] Die Gattung gilt seitdem als monotypisch.[12][19]
In seiner Erstbeschreibung ordnete Cope seine neue Gattung lediglich den Unpaarhufern zu, sah aber engere Beziehungen zu Palaeosyops, einem aus heutiger Sicht urtümlichen Vertreter der Brontotheriidae (von Cope aber noch unter der Synonymbezeichnung Limnohyus geführt). In den Folgejahren stellte Cope seine neue Gattung ausführlicher vor,[25] darunter in seinem umfassenden Werk The Vertebrata of the Tertiary Formations of the West aus dem Jahr 1884.[6] In dieser frühen Zeit ordnete er Lambdotherium weitgehend den Chalicotheriidae zu, in welchen er auch die frühen Brontotherien einsortierte.[26] Im Jahr 1889 etablierte Cope dann die eigenständige Familie der Lambdotheriidae.[27] Henry Fairfield Osborn vereinte nachfolgend Lambdotherium mit den frühen Brontotherien (die er generell als „Titanotherien“ bezeichnete).[7] Ausführlich wurde dies von ihm in seinem Werk The titanotheres of ancient Wyoming, Dakota, and Nebraska aus dem Jahr 1929 erörtert. Hierin versuchte er auch erstmals eine Schädelrekonstruktion, die sich aus mehreren unvollständigen Fossilfunden zusammensetzte.[3] Die Verbindung von Lambdotherium mit den (frühen) Brontotherien wurde nachfolgend kaum angezweifelt, wobei es hinsichtlich des Status der Lambdotherien als eigenständige Familie oder als Unterfamilie innerhalb der Brontotherien Diskussion gab.[28][20][29]
Anfang der 1980er Jahre wurde allerdings in einer unveröffentlichten Graduiertenarbeit eine engere Beziehung zwischen Lambdotherium und den Palaeotheriidae postuliert, letztere stellen Nahverwandte der Pferde dar, die aber hauptsächlich aus dem eurasischen Bereich bekannt sind. Die Schlussfolgerung basierte auf einzelnen Zahnmerkmalen, unter anderem das W-förmige Ectoloph, das auch bei einigen entwickelten Palaeotherien auftritt. Einige Autoren wie etwa Spencer George Lucas übernahmen diese Ansicht und gliederten demzufolge Lambdotherium aus den Brontotherien aus.[1] Andere Wissenschaftler blieben dem gegenüber skeptisch,[4][21] auch fielen einzelne nachfolgende phylogenetische Untersuchungen ambivalent aus, da sie sowohl eine nähere Beziehung von Lambdotherium zu den Brontotherien als auch eine Bindung des gesamten Brontotherien-Komplexes zu den Palaeotherien und Pferden unterstützten. In vielen Fällen erwiesen sich aber die Datengrundlagen als nicht ausreichend, da häufig nur die Basalformen der jeweiligen Gruppen einbezogen waren.[30][31][32] Ebenso erbrachten weiterführende Analysen etwa zum internen Aufbau der Zähne in diesem Zusammenhang keine zufriedenstellenden Ergebnisse bezüglich der genauen Verwandtschaftsverhältnisse.[33] Dies änderte sich erst mit der Entdeckung einzelner Lambdotherien-artiger Formen wie etwa Danjiangia aus Ostasien und der Einbeziehung von weitaus mehr Vertretern der Brontotherien in die Merkmalsanalysen. Dadurch lassen sich die Lambdotherien nach heutigem Stand als eine basale Entwicklungslinie der Brontotherien im weiteren Sinne auffassen.[19][22][23]
Literatur
- Ygnacio Bonillas: The Dentition of Lambdotherium. Journal of Mammalogy 17 (2), 1936, S. 139–142
- Spencer G. Lucas und Luke T. Holbrook: The skull of the Eocene perissodactyl Lambdotherium and its phylogenetic significance. New Mexico Museum of Natural History and Science Bulletin 26, 2004, S. 81–88
Einzelnachweise
- ↑ a b c Spencer G. Lucas und Luke T. Holbrook: The skull of the Eocene perissodactyl Lambdotherium and its phylogenetic significance. New Mexico Museum of Natural History and Science Bulletin 26, 2004, S. 81–88
- ↑ a b c d Ygnacio Bonillas: The Dentition of Lambdotherium. Journal of Mammalogy 17 (2), 1936, S. 139–142
- ↑ a b c d e f Henry Fairfield Osborn: The titanotheres of ancient Wyoming, Dakota, and Nebraska. United States Geological Survey Monographs 55, 1929, S. 1–894 (S. 279–288, 590–592), ([1])
- ↑ a b Bryn J. Mader: Brontotheriidae: A systematic revision and preliminary phylogeny of North American genera. In: Donald R. Prothero und Robert M. Schoch (Hrsg.): The evolution of perissodactyls. New York und London, 1989, S. 458–484
- ↑ Bryn R. Mader: A species-level revision of the North American brontotheres Eotitanops and Palaeosyops (Mammalia, Perissodactyla). Zootaxa 2339, 2010, S. 1–43
- ↑ a b Edward Drinker Cope: The Vertebrata of the Tertiary Formations of the West. Book 1. Report of the United States Geological Survey of the Territories, Washington, DC, 1884, S. 1–1009 (S. 709–712) ([2])
- ↑ a b c Henry Fairfield Osborn: Lower Eocene Titanotheres. Genera Lambdotherium, Eotitanops. Bulletin of the American Museum of Natural History 32, 1913, S. 407–415
- ↑ a b Daniel A. Guthrie: The mammalian fauna of the Lost Cabin member, Wind River formation (lower Eocene) of Wyoming. Annals of the Carnegie Museum 43, 1971, S. 47–114 ([3])
- ↑ a b Frederic Brewster Loomis: The origin of the Wasatch deposits. American Journal of Science 23 (137), 1907, S. 356–364
- ↑ Gregg F. Gunnell, William S. Bartels, Philip D. Gingerich und Victor Torres: Wapiti Valley faunas: Early and Middle Eocene fossil vertebrates from the North Fork of the Shoshone River, Park County, Wyoming. Contributions from the Museum of Paleontology, The University of Michigan 28 (1), 1992, S. 247–287
- ↑ C. Lewis Gazin: The lower Eocene Knight Formation of western Wyoming and its mammalian fauna. Smithsonian Miscellaneous Collections 117 (18), 1952, S. 1–82
- ↑ a b John-Paul Zonneveld, Gregg F. Gunnell, William S. Bartel: Early Eocene fossil vertebrates from the Southwestern Green River Basin, Lincoln and Uinta Counties, Wyoming. Journal of Vertebrate Paleontology 20 (2), 2000, S. 369–386
- ↑ Gregg F. Gunnell, John-Paul Zonneveld und William S. Bartels: Stratigraphy, mammalian paleontology, paleoecology, and age correlation of the Wasatch Formation, Fossil Butte National Monument, Wyoming. Journal of Paleontology 90 (5), 2016, S. 981–1011, doi:10.1017/jpa.2016.100
- ↑ a b Lance Grande: The lost world of Fossil Lake. Snapshot from deep time. University of Chicago Press, Chicago/London 2013, ISBN 978-0-226-92296-6, S. 1–425 (S. 270–273)
- ↑ a b Henry Fairfield Osborn: New titanotheres of Huerfano. Bulletin of the American Museum of Natural History 41, 1919, S. 557–569 ([4])
- ↑ Peter Robinson: Fossil mammals of the Huerfano Formation, Eocene of Colorado. Yale University Peabody Museum of Natural History Bulletin 21, 1966, S. 1–95
- ↑ Walter Granger: Tertiary faunal horizons in the Wind River Basin, Wyoming, with descriptions of new Eocene mammals. Bulletin of the American Museum of Natural History 28, 1910, S. 235–251
- ↑ Walter Granger: On the names of Lower Eocene faunal horizons of New Mexico and Wyoming. Bulletin of the American Museum of Natural History 33, 1914, S. 201–207
- ↑ a b c d Matthew C. Mihlbachler: Species taxonomy, phylogeny, and biogeography of the Brontotheriidae (Mammalia: Perissodactyla). Bulletin of the American Museum of Natural History 311, 2008, ISSN 0003-0090, S. 1–475
- ↑ a b Donald R. Prothero und Robert M. Schoch: Classification of the Perissodactyla. In: Donald R. Prothero und R. M. Schoch (Hrsg.): The evolution of the Perissodactyls. New York 1989, S. 530–537
- ↑ a b Kenneth D. Rose: The beginning of the age of mammls. The Johns Hopkins University Press, Baltimore, 2006, S. 1–428 (S. 250–252)
- ↑ a b Donald R. Prothero: Evolutionary Transitions in the Fossil Record of Terrestrial Hoofed Mammals. Evo Edu Outreach 2, 2009, S. 289–302, doi:10.1007/s12052-009-0136-1
- ↑ a b Bin Bai, Yuan-Qing Wang und Jin Meng: The divergence and dispersal of early perissodactyls as evidenced by early Eocene equids from Asia. Communications Biology 1, 2018, S. 115 doi:10.1038/s42003-018-0116-5
- ↑ Edward Drinker Cope: The badlands of the Wind River and their fauna. The American Naturalist 14, 1880, S. 745–748 ([5])
- ↑ Edward Drinker Cope: On the Vertebrata of the Wind River Eocene beds of Wyoming. Bulletin of the United States Geological and Geographical Survey of the Territories 6, 1881, S. 183–202 ([6])
- ↑ Edward Drinker Cope: The Perissodactyla. The American Naturalist 21, 1887, S. 985–1007 und 1060–1076 ([7])
- ↑ Edward Drinker Cope: The Vertebrata of the Swift Current River. The American Naturalist 23 (2), 1889, S. 151–155 ([8])
- ↑ George Gaylord Simpson: The principles of classification and a classification of mammals. Bulletin of the American Museum of Natural History 85, 1945, S. 1–350 (S. 137)
- ↑ Malcolm C. McKenna und Susan K. Bell: Classification of mammals above the species level. Columbia University Press, New York, 1997, S. 1–631 (S. 474)
- ↑ K. Christopher Beard: East of Eden: Asia as an important center of taxonomic origination in mammalian evolution. In K. C. Beard und M. R. Dawson (Hrsg.): Dawn of the age of mammals in Asia. Bulletin of the Carnegie Museum of Natural History 34, 1998, S. 5–39
- ↑ David J. Froehlich: Phylogenetic systematics of basal perissodactyls. Journal of Vertebrate Paleontology 19, 1999, S. 140–159
- ↑ Jerry J. Hooker und Demberelyin Dashzeveg: Evidence for direct mammalian faunal interchange between Europe and Asia near the Paleocene-Eocene boundary. In S. L. Wing, P. D. Gingerich, B. Schmitz und E. Thomas (Hrsg.): Causes and consequences of globally warm climates in the early Paleogene. Special Paper of the Geological Society of America 369, 2003, S. 479–500
- ↑ Wighart von Koenigswald, Luke T. Holbrook und Kenneth D. Rose: Diversity and Evolution of Hunter-Schreger Band Configuration in Tooth Enamel of Perissodactyl Mammals. Acta Palaeontologica Polonica 56 (1), 2011, S. 11–32, doi:10.4202/app.2010.0021